

Vorgelegt in den Sitzungen vom 8. Februar und 8. März 1865
Gedruckt in den Verhandlungen des naturforschenden Vereins in Brünn. IV. Band. Abhandlungen 1865, Brünn, 1866. Im Verlage des Verein. S. 3-47.
Künstliche Befruchtungen, welche an Zierpflanzen deshalb vorgenommen wurden, um neue Farbvarianten zu erzielen, waren die Veranlassung zu den Versuchen, die hier besprochen werden sollen. Die auffallende Regelmässigkeit, mit welcher dieselben Hybridformen immer wiederkehrten, so oft die Befruchtung zwischen gleichen Arten geschah, gab die Anregung zu weiteren Experimenten, deren Aufgabe es war, die Entwicklung der Hybriden in ihren Nachkommen zu verfolgen.
Dieser Aufgabe haben sorgfältige Beobachter, wie Kölreuter, Gärtner, Herbert, Lecocq, Wichura u. A. einen Theil ihres Lebens mit unermüdlicher Ausdauer geopfert. Namentlich hat Gärtner in seinem Werke "Die Bastarderzeugung im Pflanzenreiche" sehr schätzbare Beobachtungen niedergelegt, und in neuester Zeit wurden von Wichura gründliche Untersuchungen über die Bastarde der Weiden veröffentlicht. Wenn es noch nicht gelungen ist, ein allgemein gültiges Gesetz für die Bildung und Entwicklung der Hybriden aufzustellen, so kann das Niemanden Wunder nehmen, der den Umfang der Aufgabe kennt, und die Schwierigkeiten zu würdigen weiss, mit denen Versuche dieser Art zu kämpfen haben. Eine endgültige Endscheidung kann erst dann erfolgen, wenn Detailversuche aus den den verschiedensten Pflanzenfamilien vorliegen. Wer die Arbeiten auf diesem Gebiete überblickt, wird zu der Ueberzeugung gelangen, dass unter den zahlreichen Vesuchen keiner in dem Umfange und in der Weise durchgeführt ist, dass es möglich wäre, die Anzahl der verschiedenen Formen zu bestimmen, unter welchen die Nachkommen der Hybriden auftreten, dass man diese Formen mit Sicherheit in den einzelnen Generationen ordnen und die gegenseitigen numerischen Verhältnisse feststellen könnte. Es gehört allerdings einiger Muth dazu, sich einer so weit reichenden Arbeit zu unterziehen; indessen scheint es der einzig richtige Weg zu sein, auf dem endlich die Lösung einer Frage erreicht werden kann, welche für die Entwicklungsgeschichte der organischen Formen von nicht zu unterschützender Bedeutung ist.
Die vorliegende Abhandlung bespricht die Probe eines solches Detailversuches. Derselbe wurde sachgemäss auf eine kleine Pflanzengruppe beschränkt und ist nun nach Verlauf von acht Jahren im Wesentlichen abgeschlossen. Ob der Plan, nach welchem die einzelnen Experimente geordnet und durchgeführt wurden, der gestellten Aufgabe entspricht, darüber möge eine wohlwollende Beurtheilung entscheiden.
Der Werth und die Geltung eines jeden Experimentes wird durch die Tauglichkeit der dazu benützen Hilfsmittel, sowie durch die zweckmässige Anwendung derselben bedingt. Auch in dem vorliegenden Falle kann es nicht gleichgülitg sein, welche Pflanzenarten als Träger der Versuche gewählt und in welcher Weis diese durchgeführt wurden.
Die Auswahl der Pflanzengruppe, welche für Versuche dieser Art dienen soll, muss mit möglichster Vorsicht geschehen, wenn man nicht im Vorherein allen Erfolg in Frage stellen will.
Die Versuchspflanzen müssen nothwendig
- 1. Constant differirende Merkmale besitzen.
- Die Hybriden derselben müssen während der Blüthezeit vor der Einwirkung jedes fremdartigen Pollens geschützt sein oder leicht geschützt werden können.
- Dürfen die Hybriden und ihre Nachkommen in den aufeinander folgenden Generationen keine merkliche Störung in der Fruchtbarkeit erleiden.
Fälschungen durch fremden Pollen, wenn solche im Verlaufe des Versuches vorkämen und nicht erkannt würden, müssten zu ganz irrigen Ansichten führen. Verminderte Fruchtbarkeit, oder gänzliche Sterilität einzelner Formen, wie sie unter den Nachkommen vieler Hybriden auftreten, würden die Versuche sehr erschweren oder ganz vereiteln. Um die Beziehungen zu erkennen, in welchen die Hybridformen zu einander selbst und zu ihren Stammarten stehen, erscheint es als nothwendig, daß die Glieder der Entwicklungsreihe in der einzelnen Generation vollzählig der Beobachtung unterzogen werden.
Eine besondere Aufmerksamkeit würde gleich Anfangs den Leguminosen wegen ihres eigenthümlichen Blüthenbaues zugewendet. Versuche, welche mit mehreren Gliedern dieser Familie angestellt wurden, führten zu dem Resultate, dass das Genus Pisum den gestellten Anforderungen hinreichend entspreche. Einige ganz selbstständige Formen aus diesem Geschlechte besitzen constante, leicht und sicher zu unterscheidende Merkmale, und gegen bei gegenseitiger Kreuzung in ihren Hybriden vollkommen fruchtbare Nachkommen. Auch kann eine Störung durch fremde Pollen nicht leicht eintreten, da die Befruchtungsorgane vom Schiffchen enge umschlossen sind und die Antheren schon in der Knospe platzen, wodurch die Narbe noch vor dem Aufblühen mit Pollen überdeckt wird. Dieser Umstand ist von besonderer Wichtigkeit. Als weitere Vorzüge verdienen noch die Erwähnung die leichte Cultur dieser Pflanze im freien Lande und in Töpfen, sowie die verhältnissmassig kurze Vegetationsdauer derselben. Die künstliche Befruchtung ist allerdings etwas umständlich, gelingt jedoch fast immer. Zu diesem Zwecke wird die noch nicht vollkommen entkommen entwickelte Knospe geöffnet, das Schiffchen entfernt und jeder Staubfaden mittelst einer Pincette behutsam herausgenommen, worauf dann die Narbe sogleich mit dem fremden Pollen belegt werden kann.
Aus mehreren Samenhandlungen wurden im Ganzen 34 mehr oder weniger verschiedene Erbesensorten bezogen und einer zweijährigen Probe unterworfen. Bei einer Sorte wurden unter einer grösseren Anzahl gleicher Pflanzen einige bedeutend abweichende Formen bemerkt. Diese variirten jedoch im nächsten Jahre nicht und stimmten mit einer anderen aus derselben Samenhandlung bezogenen Art vollständig überein; ohne Zweifel waren die Samen bloss zufällig beigemengt. Alle anderen Sorten gaben durchaus gleiche und constante Nachkommen, in den beiden Probejahren wenigstens war eine wesentliche Abänderung nicht zu bemerken. Für die Befruchtung wurden 22 davon ausgewählt und jährlich, während der ganzen Versuchsdauer angebaut. Sie bewährten sich ohne alle Ausnahme.
Die systematische Einreihung derselben ist schwierig und unsicher. Wollte man die schärfste Bestimmung des Artbegriffes in Anwendung bringen, nach welcher zu einer Art nur jene Individuen gehören, die unter völlig gleichen Verhältnissen auch völlig gleiche Merkmale zeigen, so könnten nicht zwei davon zu einer Art gezählt werden. Nach der Meinung der Fachgelehrten indessen gehört die Mehrzahl der Species Pisum sativum an, während die übrigen bald als Unterarten von P. sativum, bald als selbstständige Arten angesehen und beschrieben würden, wie P. quadratum, P. saccharatum, P. umbellatum. Uebrigens bleibt die Randordnung, welche man denselben im Systeme giebt, für die in Rede stehenden Versuche völlig gleichgültig. So wenig man eine scharfe Unterscheidungslinie zwischen Species und Varietäten zu ziehen vermag, ebenso wenig ist es bis jetzt gelungen, einen gründlichen Unterschied zwischen den Hybriden der Species und Varietäten aufzustellen.
Werden zwei Pflanzen, welche in einem oder mehreren Merkmalen constant verschieden sind, durch Befruchtung verbunden, so gehen, wie zahlreiche Versuche beweisen, die gemeinsamen Merkmale unverändert auf die Hybriden und ihre Nachkommen über; je zwei differirende hingegen vereinigen sich an der Hybride zu einem neuen Merkmale, welches gewöhnlich an den Nachkommen denselben Veränderungen unterworfen ist. Diese Veränderungen für je zwei differirende Merkmale zu beobachten und das Gesetz zu ermitteln, nach welchem dieselben in den aufeinander folgenden Generationen eintreten, war die Aufgabe des Versuches. Derselbe zerfällt daher in ebenso viele einzelne Experimente, als constant differirende Merkmale an den Versuchspflanzen vorkommen.
Die verschiedenen, zur Befruchtung ausgewählten Erbsenformen zeigten Unterschiede in der Länge und Färbung des Stengels, in der Grösse und Gestalt der Blätter, in der Stellung, Farbe und Grösse der Blüthen, in der Länge der Blüthenstiele, in der Farbe, Gestalt und Grösse der Hülsen, in der Gestalt und Grösse der Samen, in der Färbung der Samenschale und des Albumens. Ein Theil der angeführten Merkmale lässt jedoch eine sichere und scharfe Trennung nicht zu, indem der Unterschied auf einem oft schwierig zu bestimmenden "mehr oder weniger" beruht. Solche Merkmale waren für die Einzelversuche nicht verwendbar, diese konnten sich nur auf Charaktere beschränken, die an den Pflanzen deutlich und entschieden hervortreten. Der Erfolg musste entlich zeigen, ob sie in hybrider Vereinigung sämmtlich ein übereinstimmendes Verhalten beobachten, und ob daraus auch ein Urtheil über jene Merkmale möglich wird, welche eine untergeordnete typische Bedeutung haben.
Die Merkmale, welche in die Versuche aufgenomen wurden, beziehen sich:
1. auf den Unterschied in der Gestalt der reifen Samen. Diese sind entweger kugelrund oder rundlich, die Einsenkungen, welche an der Oberfläche vorkommen, immer nur seicht, oder sie sind unregelmässig kantig, tief runzelig (P. quadratum);In zwei von den angeführten differirenden Merkmalen wurden durch Befruchtung vereinigt. Für den2. auf den Unterschied in der Färbung des Samenalbumens (Endosperm's). Das Albumen der reifen Samen ist entweder blassgelb, hellgelb oder orange gefärbt, oder es besitzt eine mehr oder weniger intensiv grüne Farbe. Dieser Farbenunterschied ist an den Samen deutlich zu erkennen, da ihre Schalen durchscheinend sind;
3. auf den Unterschied in der Färbung der Samenschale. Diese ist entweder weiss gefärbt, womit auch constant die weisse Blüthenfarbe verbunden ist, oder sie ist grau, graubraun, lederbraun mit oder ohne violetter Punctirung, dann erscheint die Farbe der Fahne violett, die der Flügel purpurn, und der Stengel an den Blattachseln röthlich gezeichnet. Die grauen Samenschalen werden im kochenden Wasser schwarzbraun;
4. auf den Unterschied in der Form der reifen Hülse. Diese ist entweder einfach gewölbt, nie stellenweise verengt, oder sie ist zwischen den Samen tief eingeschnürt und mehr oder weniger runzelig (P. saccharatum).
5. auf den Unterschied in der Farbe der unreifen Hülse. Sie ist entweder licht- bis dunkelgrün oder lebhaft gelb gefärbt, an welcher Färbung auch Stengel, Blattrippen und Kelch theilnehmen *{Fußnote: Eine Art besitzt eine schöne braunrothe Hülsenfarbe, welche gegen die Zeit der Reife hin in Violett und Blau übergeht. Der Versuch über dieses Merkmal wurde erst im verflossenen Jahre begonnen.};
6. auf den Unterschied in der Stellung der Blüthen. Sie sind entweder axenständig, d. i längs der Axe vertheilt, oder sie sind entständig, am Ende der Axe gehäuft und fast in eine kurze Trugdolde gestellt; dabei ist der obere Theil des Stengels im Querschnitte mehr oder weniger erweitert (P. umbellatum);
7. auf den Unterschied in der Axenlänge. Die Länge der Axe ist bei den einzelnen Formen sehr verschieden, jedoch für jede insofern ein constantes Merkmal, als dieselbe bei gesunden Pflanzen, die in gleichem Boden gezogen werden, nur unbedeutenden Aenderungen unterliegt. Bei den Versuchen über dieses Merkmal wurde der sicheren Unterscheidung wegen stets die lange Axe von 6-7' mit der kurzen von 3/4' bis 1 1/2' verbunden.
1. Vers. wurden 60 Befrucht. an 15 Pflanzen vorgenommenVon einer grösseren Anzahl Pflanzen derselben Art wurden zur Befruchtung nur die kräftigsten ausgewählt. Schwache Exemplare geben immer unsichere Resultate, weil schon in der ersten Generation der Hybriden und noch mehr in der folgenden manche Abkömmliche entweder gar nicht zur Blüthe gelangen, oder doch wenige und schlechte Samen bilden.2. Vers. wurden 58 Befrucht. an 10 Pflanzen vorgenommen
3. Vers. wurden 35 Befrucht. an 10 Pflanzen vorgenommen
4. Vers. wurden 40 Befrucht. an 10 Pflanzen vorgenommen
5. Vers. wurden 23 Befrucht. an 5 Pflanzen vorgenommen
6. Vers. wurden 34 Befrucht. an 10 Pflanzen vorgenommen
7. Vers. wurden 37 Befrucht. an 10 Pflanzen vorgenommen.
Ferner wurde bei sämmtlichen Versuchen die wechselseitige Kreuzung durchgeführt, in der Weise nämlich, dass jene der beiden Arten, welche bei einer Anzahl Befruchtungen als Samenpflanze diente, bei der anderen als Pollenpflanze verwendet wurde.
Die Pflanzen wurden auf Gartenbeeten, ein kleiner Theil in Töpfen gezogen, und mittelst Stäben, Baumzweigen und gespannten Schnüren in der natürlichen aufrechten Stellung erhalten. Für jeden Versuch wurde eine Anzahl Topfpflanzen während der Blüthezeit in ein Gewächshaus gestellt, sie sollten für den Hauptversuch im Garten als Controlle dienen bezüglich möglicher Störungen durch Insecten. Unter jenen, welche die Erbsenpflanze besuchen, könnte die Käferspecies Bruchus pisi dem Versuche gefährlich werden, falls sie in grösserer Menge erscheint. Das Weibchen dieser Art legt bekanntlich seine Eier in die Blüthe und öffnet dabei das Schiffchen; an den Tarsen eines Exemplares, welches in einer Blüthe gefangen wurde, konnten unter der Lupe deutlich einige Pollenzellen bemerkt werden. Es muss hier noch eines Umstandes Erwähnung geschehen, der möglicher Weise die Einmengung fremden Pollens veranlassen könnte. Es kommt nämlich in einzelnen seltenen Fällen vor, dass gewisse Theile der übrigens ganz normal entwickelten Blüthe verkümmern, wodurch eine theilweise Entblössung der Befruchtungsorgane herbeigeführt wird. So wurde eine mangelhafte Entwicklung des Schiffchens beobachtet, wobei Griffel und Antheren zum Theile unbedeckt blieben. Auch geschieht es bisweilen, dass der Pollen nicht zur vollen Ausbildung gelangt. In diesem Falle findet während des Blühens eine allmähliche Verlängerung des Griffels statt, bis die Narbe an der Spitze des Schiffchens hervortritt. Diese merkwürdige Erscheinung wurde auch an Hybriden von Phaseolus und Lathyrus beobachtet.
Die Gefahr einer Fälschung durch den fremden Pollen ist jedoch bei Pisum eine sehr geringe und vermag keineswegs das Resultat im Grossen und Ganzen zu stören. Unter mehr als 10.000 Pflanzen, welche genauer untersucht wurden, kam der Fall nur einige wenige Male vor, dass eine Einmengung nicht zu bezweifeln war. Da im Gewächshaus niemals eine solche Störung beobachtet wurde, liegt wohl die Vermuthung nahe, dass Bruchus pisi und vielleicht auch die angeführten Abnormitäten im Blüthenbau die Schuld daran tragen.
Schon die Versuche, welche in früheren Jahren an Zierpflanzen vorgenommen wurden, lieferten den Beweis, dass die Hybriden in der Regel nicht die genaue Mittelform zwischen den Stammarten darstellen. Bei einzelen mehr in die Augen springenden Merkmalen, wie bei solchen, die sich auf die Gestalt und Grösse der Blätter, auf die Behaarung der einzelnen Theile u. s. w. beziehen, wird in der That die Mittelbildung fast immer ersichtlich; in anderen Fällen hingegen besitzt das eine der beiden Stamm-Merkmale ein so grosses Uebergewicht, dass es schwierig oder ganz unmöglich ist, das andere an der Hybride aufzufinden.
Ebenso verhält es sich mit den Hybriden bei Pisum. Jedes von den 7 Hybridenmerkmalen gleicht dem einen der beiden Stamm-Merkmale entweder so vollkommen, dass das andere der Beobachtung entschwindet, oder ist demselben so ähnlich, dass eine sichere Unterscheidung nicht stattfinden kann. Dieser Umstand ist von grosser Wichtigkeit für die Bestimmung und Einreihung der Formen, unter welchen die Nachkommen der Hybriden erscheinen. In der weiteren Besprechung werden jene Merkmale, welche ganz oder fast unverändert in die Hybride-Verbindung übergehen, somit selbst die Hybridenmerkmale repräsentiren, als dominante und jene, welche in der Verbindung latent werden, als recessive bezeichnet. Der Ausdruck "recessiv" wurde deshalb gewählt, weil die damit benannten Merkmale an den Hybriden zurücktreten oder ganz verschwinden, jedoch unter den Nachkommen derselben, wie später gezeigt wird, wieder unverändert zum Vorschein kommen.
Es wurde ferner durch sämmtliche Versuche erwiesen, dass es völlig gleichgültig ist, ob das dominirende Merkmal der Samen- oder der Pollenpflanze angehört; die Hybridform bleibt in beiden Fällen genau dieselbe. Diese interessante Erscheinung wird auch von Gärtner hervorgehoben, mit dem Bemerken, dass selbst der geübteste Kenner nicht im Stande ist, an einer Hybride zu unterscheiden, welche von den beiden verbundenen Arten die Samen- oder Pollenpflanze war.
Von den differirenden Merkmalen, welche in die Versuche eingeführt wurden, snd nachfolgende dominirend:
1. die runde oder rundliche Samenform mit oder ohne seichte Einsenkungen;Was das letztere Merkmal angeblangt, muss bemerkt werden, dass die längere der beiden Stammaxen von der Hybride gewöhnlich noch übertroffen wird, was vielleicht nur der grossen Ueppigkeit zuzuschrieben ist, welche in allen Pflanzentheilen auftritt, wenn Axen von sehr verschiedener Länge verbunden sind. So z. B. gaben bei wiederholtem Versuche Axen von 1' und 6' Länge in hybrider Vereinigung ohne Ausnahmen Axen, deren Länge zwischen 6- 71/2' schwankte. Die Hybriden der Samenschale sind öfters mehr punctirt, auch fliessen die Punkte bisweilen in kleinere bläulich-violette Flecke zusammen. Die Punctirung erscheint häufig auch dann, wenn sie selbst dem Stamm-Merkmale fehlt.2. die gelbe Färbung des Samenalbumens;
3. die graue, graubraune oder lederbraune Farbe der Samenschale, in Verbindung mit violettrother Blüthe und röthlicher Makel in den Blattachseln;
4. die einfach gewölbte Form der Hülse;
5. die grüne Färbung der unreifen Hülse, in Verbindung mit der gleichen Farbe des Stengels, der Blattrippen und des Kelches;
6. die Vertheilung der Blüthen längs des Stengels;
7. das Längenmaass der grösseren Axe.
Die Hybridformen der Samengestalt und des Albumens entwickeln sich unmittelbar nach der künstlichen Befruchtung durch die blosse Einwirkung des fremden Pollens. Sie können daher schon im ersten Versuchsjahre beobachtet werden, während alle übrigen selbstverständlich erst im folgenden Jahre an jenen Pflanzen hervortreten, welche aus den befruchteten Samen gezogen werden.
In dieser Generation treten nebst den dominirenden Merkmalen auch die recessiven in ihrer vollen Eigenschaft wieder auf, und zwar in dem entschieden ausgesprochenen Durchschnittsverhältnisse 3:1, so dass unter je vier Pflanzen aus dieser Generation drei den dominirenden und eine den recessiven Charakter erhalten. Es gilt das ohne Ausnahme für alle Merkmale, welche in die Versuche aufgenommen waren. Die kantig runzelige Gestalt der Samen, die grüne Färbung des Albumens, die weisse Farbe der Samenschale und der Blüthe, die Einschnürungen an den Hülsen, die gelbe Farbe der unreifen Hülse, des Stengelns,, Kelches und der Blattrippen, der trugdoldenförmige Blüthenstand und die zwergartige Axe kommen in den angeführten numerischen Verhältnisse wiedre zum Vorschein ohne irgend eine wesentliche Abänderung. Uebergangsformen wurden bei keinem Versuche beobachtet.
Da die Hybriden, welche aus wechselseitiger Kreuzung hervorgingen, eine völlig gleiche Gestalt besassen und auch in ihrer Weiterentwicklung keine bemerkenswerthe Abweichung ersichtlich wurde, konnten die beiderseitigen Resultate für jeden Versuch unter eine Rechnung gebracht werden. Die Verhältnisszahlen, welche für je zwei differirende Merkmale gewonnen wurden, sind folgende:
1. Versuch. Gestalt der Samen. 253 Hybriden wurden im zweiten Versuchsjahre 7324 Samen erhalten. Derunter waren rund oder rundlich 5475, und kantig runzelig 1850 Samen. Daraus ergiebt sich das Verhältniss 2,96 : 1.Bei diesen beiden Versuchen erhält man gewöhnlich aus jeder Hülse beiderlei Samen. Bei gut ausgebildeten Hülsen, welche durchschnittlich 6-9 Samen enthielten, kam es öfters vor, dass sämmtliche Samen rund (Versuch 1) oder sämmtliche gelb (Versuch 2) waren; hingegen wurden mehr als 5 kantige oder 5 grüne in einer Hülse niemals beobachtet. Es scheint keinen Unterschied zu machen, ob die Hülse sich früher oder später an der Hybride entwickelt, ob sie der Hauptaxe oder einer Nebenaxe angehört. An einigen wenigen Pflanzen kamen in den zuerst gebildeten Hülsen nur einzelne Samen zur Entwicklung, und diese besassen dann ausschliesslich das eine der beiden Merkmale; in den später gebildeten Hülsen blieb jedoch das Verhältniss normal. So wie in einzelnen Hülsen, ebenso variit die Vertheilung der Merkmale auch bei einzelnen Pflanzen. Zur Veranschaulichung mögen die ersten 10 Glieder aus beiden Versuchsreihen dienen:2. Versuch. Färbung des Albumens. 258 Pflanzen gaben 8023 Samen, 6022 gelbe und 2001 grüne; daher stehen jene zu diesen im Verhältnisse 3,01 : 1.
| Pflanze | rund | kantig | ||
Als Extrem in der Vertheilung der beiden Samenmerkmale an einer Pflanze wurde beobachtet bei dem 1. Versuche 43 runde und nur 2 kantige, ferner 14 runde und 15 kantige Samen. Bei dem 2. Versuche 32 gelbe und nur 1 grüner Samen, aber auch 20 gelbe und 19 grüne.
Diese beiden Versuche sind wichtig für die Feststellung der mittleren Verhältnisszahlen, weil sie bei einer geringeren Anzahl von Versuchspflanzen sehr bedeutende Durchschnitte möglich machen. Bei der Abzählung der Samen wird jedoch, namentlich beim 2. Versuche, einige Aufmerksamkeit erfordert, da bei einzelnen Samen mancher Pflanzen die grüne Färbung des Albumens weniger entwickelt wird und anfänglich leicht übersehen werden kann. Die Ursache des theilweisen Verschwindens der grünen Färbung steht mit dem Hybriden-Charakter der Pflanzen in keinem Zusammenhange, indem dasselbe an der Stammpflanze ebenfalls vorkommt; auch beschränkt sich diese Eigenthümlichkiet nur auf das Individuum und vererbt sich nicht auf die Nachkommen. An luxurirenden Pflanzen wurde diese Erscheinung öfter beobachtet. Samen, welche während ihrer Entwicklung von Insecten beschädigt wurden, variiren oft in Farbe und Gestalt, jedoch sind bei einiger Uebung im Sortiren Fehler leicht zu vermeiden. Es ist fast überflüssig zu erwähnen, dass die Hülsen so lange an der Pflanze bleiben müssen, bis sie vollkommen ausgereift und trocken geworden sind, weil erst dann die Gestalt und Färbung der Samen vollständig entwickelt ist.
3. Versuch. Farbe der Samenschale. Unter 929 Pflanzen brauchen 705 violettrothe Blüthen und graubraune Samenschalen; 224 hatten weisse Blüthen und weiss Samenschalen. Daraus ergiebt sich das Verhältniss 3,15 : 1.Werden die Resultate sämmtlicher Versuche zusammengefasst, so ergibt sich zwischen der Anzahl der Formen mit dem dominirenden und recessiven Merkmale das Durchschnittsverhältniss 2,98 : 1 oder 3 : 1.4. Versuch. Gestalt der Hülsen. Von 1181 Pflanzen hatten 882 einfach gewölbte, 299 eingeschnürte Hülsen. Daher das Verhältniss 2,95 : 1.
5. Versuch. Färbung der unreifen Hülse. Die Zahl der Versuchspflanzen betrug 580, wovon 428 grüne und 152 gelbe Hülsen besassen. Daher stehen jene zu diesen in dem Verhältnisse 2,82 : 1. p align="justify"> 6. Versuch. Stellung der Blüthen. Unter 858 Fällen waren die Blüthen 651 Mal axenständig und 207 Mal endständig. Daraus das Verhältnis 3,14 : 1.
7. Versuch. Länge der Axe. Von 1064 Pflanzen hatten 787 die lange, 277 die kurze Axe. Daher das gegenseitige Verhältniss 2,84 : 1. Bei diesem Versuche wurden die zwergartigen Pflanzen behutsam ausgehoben und auf eigene Beete versetzt. Diese Vorsicht war nothwendig, weil sie sonst mitten unter ihren hochrankenden Geschwistern hätten verkümmern müssen. Sie sind schon in der ersten Jugendzeit an dem gedrungenen Wuchse und den dunkelgrünen dicken Blättern leicht zu unterscheiden.
Das dominirende Merkmal kann hier eine doppelte Bedeutung haben, nämlich die des Stammcharakters oder des Hybriden- Merkmales. In welcher von beiden Bedeutungen dasselbe in jedem einzelnen Falle vorkommt, darüber kann nur die nächste Generation entscheiden. Als Stamm-Merkmal muss dasselbe unverändret auf sämmtliche Nachkommen übergehen, als hybrides Merkmal hingegen ein gleiches Verhalten wie in der ersten Generation beobachten.
>
Jene Formen, welche in der ersten Generation den recessiven Charakter erhalten, variiren in der zweiten Generation in Bezug auf diesen Charakter nicht mehr, sie bleiben in ihren Nachkommen constant.
Anders verhält es sich mit jenen, welche in der ersten Generation das dominirende Merkmal besitzen. Von diesen geben zwei Theile Nachkommen, welche in dem Verhältnisse 3 : 1 das dominirende und recessive Merkmal an sich tragen, somit genau dasselbe Verhalten zeigen, wie die Hybridformen; nur ein Theil bleibt mit dem dominirenden Merkmale constant.
Die einzelnen Versuche lieferten nachfolgende Resultate:
1. Versuch. Unter 565 Pflanzen, welche aus runden Samen der ersten Generation gezogen wurden, brachten 193 wieder nur runde Samen und blieben demnach in diesem Merkmale constant; 372 aber gaben runde und kantige Samen zugleich, in dem Verhältnisse 3 : 1. Die Anzahl der Hybriden verhielt sich daher zu der Zahl der Constanden wie 1,93 : 1.2. Versuch. Von 519 Pflanzen, welche aus Samen gezogen wurden, deren Albumen in der ersten Generation die gelbe Färbung zeigte, gaben 166 ausschliesslich gelbe, 353 aber gelbe und grüne Samen in dem Verhältnisse 3 : 1. Es erfolgte daher eine Theilung in hybride und constante Formen nach dem Verhältnisse 2,13 : 1.
Für jeden einzelnen von den nachfolgenden Versuchen wurden 100 Pflanzen ausgewählt, welche in dre ersten Generation das dominirende Merkmal besassen, und um die Bedeutung desselben zu prüfen, von jeder 10 Samen angebaut.
3. Versuch. Die Nachkommen von 36 Pflanzen brachten ausschliesslich graubraune Samenschalen; von 64 Pflanzen wurden theils graubraune theils weisse erhalten.
4. Versuch. Die Nachkommen von 29 Pflanzen hatten nur einfach gewölbte Hülsen, von 71 hingegen theils gewölbte, theils eingeschnürte.
5. Versuch. Die Nachkommen von 40 Pflanzen hatten bloss grüne Hülsen, die von 60 Pflanzen theils grüne, theils gelbe.
6. Versuch. Die Nachkommen von 33 Pflanzen hatten bloss axenständige Blüthen, bei 67 hingegen waren sie theils axanständig, theils endständig.
7. Versuch. Die Nachkommen von 29 Pflanzen erhielten die lange Axe, die von 72 Pflanzen theils die lange, theils die kurze.
Bei jedem dieser Versuche wird eine bestimmte Anzahl Pflanzen mit dem dominirenden Merkmal constant. Für die Beurtheilung des Verhältnisses, in welchem die Ausscheidung der Formen mit dem constant bleibenden Merkmale erfolgt, sind die beiden ersten Versuche von besonderem Gewichte, weil bei diesen eine grössere Anzahl Pflanzen verglichen werden konnte. Die Verhältnisse 1,93 : 1 und 2,13 : 1 geben zusammen fast genau das Durchschnittsverhältniss 2 : 1. Der 6. Versuch hat ein ganz überdurchschnittliches Resultat, bei den anderen schwankt das Verhältniss mehr oder weniger, wie es bei der geringen Anzahl von 100 Versuchspflanzen nicht anders zu erwarten war. Der 5. Versuch, welche die grösste Abweichung zeigte, wurde wiederholt und dann, statt des Verhältnisses 60 : 40, das Verhältniss 65 : 35 erhalten. Das Durchschnittsverhältniss 2 : 1 erscheint demnach als gesichert. Es ist damit erwiesen, dass von jenen Formen, welche in der ersten Generation das dominirende Merkmal besitzen, zwei Theile den hybriden Charakter an sich tragen, ein Theil aber mit dem dominirenden Merkmale constant bleibt.
Das Verhältniss 3 : 1, nach welchem die Vertheilung des dominirenden und recessiven Charakters in der ersten Generation erfolgt, löst sich demnach für alle Versuche in die Verhältnisse 2 : 1 : 1 auf, wenn man zugleich das dominirende Merkmal in seiner Bedeutung als hybrides Merkmal und als Stammcharakter unterscheidet. Da die Glieder der ersten Generation unmittelbar aus den Samen der Hybriden hervorgehen, wird es nun ersichtlich, dass die Hybriden je zweier differirender Merkmale Samen bilden, von denen die eine Hälfte wider die Hybridform entwickelt, während die andere Pflanzen giebt, welche constant bleiben und zu gleichen Theilen den dominirenden und recessiven Charakter erhalten.
Bezweichnet A das eine der beiden constanten Merkmale, z. B. das dominirende, a das recessive, und Aa die Hybridform, in welcher beide vereinigt sind, so ergiegt der Ausdruck:
die Entwicklungsreihe für die Nachkommen der Hybriden je zweier differirender Merkmale.
Die von Gärtner, Kölreuter und Anderen gemachte Wahrnehmung, dass Hybriden die Neigung besitzen, zu den Stammarten zurückzukehren, ist auch durch die besprochenen Versuche bestätigt. Es lässt sich zeigen, dass die Zahl der Hybriden, welche aus einer Befruchtung stammen, gegen die Anzahl der constant gewordenen Formen und ihrer Nachkommen von Generation zu Generation um ein Bedeutendes zurückbleibt, ohne dass sie jedoch ganz verschwinden könnten. Nimmt man durchschnittlich für alle Pflanzen in allen Generationen eine gleich grosse Fruchtbarkeit an, erwägt man ferner, dass jede Hybride Samen bildet, aus denen zur Hälfte wieder Hybriden hervorgehen, während die andere Hälfte mit beiden Merkmalen zu gleichen Theilen constant wird, so ergeben sich die Zahlenverhältnisse für die Nachkommen in jeder Generation aus folgender Zusammenstellung, wobei A und a wieder die beiden Stamm-Merkmale und Aa die Hybridform bezeichnet. Der Kürze wegen möge die Annahme gelten, dass jede Pflanze in jeder Generation nur 4 Samen bildet.
| in Verhältniss gestellt: | |||||
| Generation | A | Aa | a | A : Aa : a | |
| 1 : 2 : 1 | |||||
| 3 : 2 : 3 | |||||
| 7 : 2 : 7 | |||||
| 15 : 2 : 15 | |||||
| 2n- 1 : 2 : 2n-1 | |||||
In der 10. Generation z. B. ist 2n-1 = 1023. Es giebt somit unter je 2048 Pflanzen, welche aus dieser Generation hervorgehen, 1023 mit dem constanten dominirenden, 1023 mit dem recessiven Merkmale und nur 2 Hybriden.
Die Nachkommen der Hybriden, inwelchen mehrere differirende Merkmale verbunden sind.
Für die eben besprochenen Versuche wurden Pflanzen verwendet, welche nur in einem wesentlichen Merkmale verschieden waren. Die nächste Aufgabe bestand darin, zu untersuchen, ob das gefundene Entwicklungsgesetz auch dann für je zwei differirende Merkmale gelte, wenn mehrere verschiedene Charaktere durch Befruchtung in der Hybride vereinigt sind.
Was die Gestalt der Hybriden in diesem Falle angelangt, zeigten die Versuche übereinstimmend, dass dieselbe stets jener der beiden Stammpflanezn näher steht, welche die grössere Anzahl von dominirenden Merkmalen besitzt. Hat z. B. die Samenpflanze eine kurze Axe, endständige weisse Blüthen und einfach gewölbte Hülsen; die Pollenpflanze hingegen eine lange Axe, axenständige violett-rothe Blüthen und eingeschnürte Hülsen, so erinnert die Hybride nur durch die Hülsenform an die Samenpflanze, in den übrigen Merkmalen stimmt sie mit der Pollenpflanze überein. Besitzt eine der beiden Stammarten nur dominirende Merkmale, dann ist die Hybride von derselben kaum oder gar nicht zu unterscheiden.
Mit einer grösseren Anzahl Pflanzen wurden zwei Versuche durchgeführt. Bei dem ersten Versuche waren die Stammpflanzen in der Gestalt der Samen und in der Färbung des Albumens verschieden; bei dem zweiten in der Gestalt der Samen, in der Färbung des Albumens und in der Farbe der Samenschale. Versuche mit Samenmerkmalen führen einfachsten und sichersten zum Ziele.
Um eine leichtere Uebersicht zu gewinnen, werden bei diesen Versuchen die differirenden Merkmale der Samenpflanze mit A, B, C, jene der Pollenpflanze mit a, b, c und die Hybridformen dieser Merkmale mit Aa, Bb, Cc bezeichnet.
Erster Versuch:
| AB Samenpflanze, | ab Pollenpflanze, |
| A Gestalt rund, | a Gestalt kantig, |
| B Albumen gelb, | b Albumen grün. |
Die befruchteten Samen erschienen rund und gelb, jenen der Samenpflanze ähnlich. Die daraus gezogenen Pflanzen gaben Samen von viererlei Art, welch oft gemeinschaftlich in einer Hülse lagen. Im Ganzen wurden von 15 Pflanzen 556 Samen erhalten, von diesen waren:
| 315 | rund und gelb, |
| 101 | kantig und gelb, |
| 108 | rund und grün |
| 32 | kantig und grün. |
Alle wurden im nächsten Jahre angebaut. Von den runden gelben Samen gingen 11 nicht auf und 3 Pflanzen kamen nicht zur Fruchtbildung. Unter den übrigen Pflanzen hatten:
| 38 runde gelbe Samen | AB |
| 65 runde gelbe und grüne Samen | ABb |
| 60 runde gelbe und kantige gelbe Samen | AaB |
| 138 runde gelbe und grüne, kantige gelbe und grüne Samen | AaBb |
Von den kantigen gelben Samen kamen 96 Pflanzen zur Fruchtbildung, wovon
| 28 nur kantige gelbe Samen hatten | aB |
| 68 kantige, gelbe und grüne Samen | aBb |
Von 108 runden grünen Samen brauchen 102 Pflanzen Früchte, davon hatten:
| 35 nur röunde grüne Samen | Ab |
| 67 runde und kantige grüne Samen | Aab |
Die kantigen grünen Samen gaben 30 Pflanzen mit durchaus gleichem Samen; sie blieben constant ab.
Die Nachkommen der Hybriden erscheinen demnach unter 9 verschiedenen Formen und zum Theile in sehr ungleicher Anzahl. Man erhält, wenn dieselben zusammengestellt und geordnet werden:
| 38 | Pflanzen mit der Bezeichnung | AB |
| 35 | Pflanzen mit der Bezeichnung | Ab |
| 28 | Pflanzen mit der Bezeichnung | aB |
| 30 | Pflanzen mit der Bezeichnung | ab |
| 65 | Pflanzen mit der Bezeichnung | ABb |
| 68 | Pflanzen mit der Bezeichnung | aBb |
| 60 | Pflanzen mit der Bezeichnung | AaB |
| 67 | Pflanzen mit der Bezeichnung | Aab |
| 138 | Pflanzen mit der Bezeichnung | AaBb |
Sämmtliche Formen lassen sich in drei wesentlich verschiedene Abtheilungen bringen. Die erste umfasst jene mit der Bezeichnung AB, Ab, aB, ab; sie besitzen nur constante Merkmale und ändern sich in den nächsten Generationen nicht mehr. Jede dieser Formen ist durchschnittlich 33 Mal vertreten. Die zweite Gruppe enthält die Formen ABb, aBb, AaB, Aab; diese sind in einem Merkmale constant, in dem anderen hydrid, und variiren in der nächsten Generation nur hinsichtlich des hybriden Merkmales. Jede davon erscheint im Durchschnitte 65 Mal. Die Form AaBb kommt 138 Mal vor, ist in beiden Merkmalen hybrid, und verhält sich genau so, wie die Hybride, von der sie abstammt.
Vergleicht man die Anzahl, in welcher die Formen dieser Abtheilungen vorkommen, so sind die Durchschnittsverhältnisse 1 : 2 : 4 nicht zu verkennen. Die Zahlen 33, 65, 138 geben ganz günstige Annäherungswerthe an die Verhältnisszahlen 33, 66 und 132.
Die Entwicklungsreihe besteht demnach aus 9 Gliedern, 4 davon kommen in derselben je einmal vor und sind in beiden Merkmalen constant; die Formen AB, ab gleichen den Stammarten, die beiden anderen stellen die ausserdem noch möglichen constanten Combinationen zwischen den verbundenen Merkmalen A, a, B, b vor. Vier Glieder kommen je 2 Mal vor und sind in einem Merkmale constant, in dem anderen hybrid. Ein Glied tritt 4 Mal auf und ist in beiden Merkmalen hybrid. Daher entwickeln sich die Nachckommen der Hybriden, wenn in denselben zweierlei differirende Merkmale verbunden sind, nach dem Ausdrucke:
Diese Entwicklungsreihe ist unbestritten eine Combinationsreihe, in welche die beiden Entwicklungsreiehn für die Merkmale A und a, B und b gliedweise verbunden sind. Man erhält die Glieder der Reihe vollzählig durch die Combinirung der Ausdrücke:
Zweiter Versuch:
| ABC | Samenpflanze | abc | Pollenpflanze |
| A | Gestalt rund, | a | Gestalt kantig, |
| B | Albumen gelb, | b | Albumen grün, |
| C | Schale graubraun, | c | Schale weiss. |
| 8 | Pflanzen | ABC |
| 14 | " | ABc |
| 9 | " | ABc |
| 11 | " | Abc |
| 22 | " | ABCc |
| 17 | " | AbCc |
| 25 | " | aBCc |
| 20 | " | abCc |
| 45 | " | ABbCc |
| 36 | " | aBbCc |
| 38 | " | AaBCc |
| 40 | " | AabCc |
| 8 | " | aBC |
| 10 | " | aBc |
| 10 | " | abC |
| 7 | " | abc |
| 15 | " | ABbC |
| 18 | " | ABbc |
| 19 | " | aBbC |
| 24 | " | aBbc |
| 14 | " | AaBC |
| 18 | " | AaBc |
| 20 | " | AabC |
| 16 | " | Aabc |
| 49 | " | AaBbC |
| 48 | " | AaBbc |
| 78 | " | AaBbCc |
Die Entwicklungsreihe unfasst 27 Glieder, davon sind 8 in allen Merkmalen constant, und jede kommt durchschnittlich 10 Mal vor; 12 sind in zwei Merkmalen constant, in dem dritten hybrid, jede erscheint im Durchschnitte 19 Mal; 6 sind in einem Merkmal constant, in den beiden anderen hybrid, jede davon tritt durchschnittlich 43 Mal auf; eine Form kommt 78 Mal vor und ist in sämmtlichen Merkmalen hybrid. Die Verhältnisse 10 : 19 : 43 : 78 kommen den Verhältnissen 10 : 20 : 40 : 80 oder 1 : 2 : 4 : 8 so nahe, dass letztere ohne Zweifel die richtigen Werthe darstellen.
Die Entwicklung der Hybriden, wenn ihre Stammarten in drei Merkmalen verschieden sind, erfolgt daher nach dem Ausdrucke:
Auch hier liegt eine Combinationsreihe vor, in welcher die Entwicklungsreihe für die Merkmale A und a, B und b, C und c mit einander verbunden sind. Die Ausdrücke:
geben sämmtliche Glieder der Reihe. Die constanten Verbindungen, welche in derselben vorkommen, entsprechen allen Combinationen, welche zwischen den Merkmalen A, B, C, a, b, c möglich sind; zwei davon ABC und abc gleichen den beiden Stammpflanzen.
Ausserdem wurden noch mehrere Experimente mit einer geringeren Anzahl Versuchspflanzen durchgeführt, bei welchen die übrigen Merkmale zu zwei und drei hybrid verbunden waren; alle lieferten annährnd gleiche Resultate. Es unterliegt daher keinem Zweifel, dass für sämmtliche in die Versuche aufgenommenen Merkmale der Satz Gültigkeit habe: die Nachkommen der Hybriden, in welchen mehrere wesentlich verschiedene Merkmale vereinigt sind, stellen die Glieder einer Combinationsreihe vor, in welchen die Entwicklungsreihen für je zwei differirende Merkmale verbunden sind. Damit ist zugleich erwiesen, dass das Verhalten je zweier differirender Merkmale in hybrider Verbindung unabhängig ist von den anderweitigen Unterschieden an den beiden Stammpflanzen.
Bezeichnet n die Anzahl der charakteristischen Unterschiede an den beiden Stammpflanzen, so giebt 3n die Gliederzahl der Combinationsreihe, 4n die Anzahl der Individuen, welche in die Reihe gehören, und 2n die Zahl der Verbindungen, welche constant bleiben. So enthält z. B. die Reieh, wenn die Stammarten in 4 Merkmalen verschieden sind, 34 = 81 Glieder, 44 = 256 Individuen und 24 = 16 constante Formen; oder was dasselbe ist, unter je 256 Nachkommen der Hybriden giebt es 81 verschiedene Verbindungen, von denen 16 constant sind.
Alle constanten Verbindungen, welche bei Pisum durch Combinirung der angeführten 7 charakteristischen Merkmale möglich sind, wurden durch wiederholte Kreuzungen auch wirklich erhalten. Ihre Zahl ist durch 27 = 128 gegeben. Damit ist zugleich der faktische Beweis geliefrt, dass constante Merkmale, welche an verschiedenen Formen einer Pflanzensippe vorkommen, auf dem Wege der wiederholten künstlichen Befruchtung in alle Verbindungen treten können, welche nach den Regeln der Combination möglich sind.
Ueber die Blüthezeit der Hybriden sind die Versuche noch nicht abgeschlossen. Soviel kann indessen schon angegeben werden, dass dieselbe fast genau in der Mitte zwischen jener der Samen- und Pollenpflanze steht, und die Entwicklung der Hybriden bezüglich dieses Merkmals wahrscheinlich in der nämlichen Weise erfolgt, wie es für die übrigen Merkmale der Fall ist. Die Formen, welche für Versuche dieser Art gewählt werden, müssen in der mittleren Blüthezeit wenigstens um 20 Tage verschieden sein; ferner ist es nothwendig, dass die Samen beim Anbaue alle gleich tief in die Erde versenkt werden, um ein gleichzeitiges Keimen zu erzielen, dass ferner während der ganzen Blüthezeit grössere Schwankungen in der Temperatur und die dadurch bewirkte theilweise Beschleunigung oder Verzögerung des Aufblühens in Rechnung gezogen werden. Man sieht, dass dieser Versuch mancherlei Schwierigkeiten zu überwinden hat und grosse Aufmerksamkeit erfordert.
Versuchen wir die gewonnenen Resultate kurz zusammenzufassen, so finden wir, dass jene differirenden Merkmale, welche an den Versuchspflanzen eine leichte und sichere Unterscheidung zulassen, in hybrider Vereinigung ein völlig übereinstimmendes Verhalten beobachten. Die Nachkommen der Hybriden je zweier differirender Merkmale sind zur Hälfte wieder Hybriden, während die andere Hälfte zu gleichen Theilen mit dem Charakter der Samen- und Pollenpflanze constant wird. Sind mehrere differirende Merkmale durch Befruchtung in einer Hybride vereinigt, so bilden die Nachkommen derselben die Glieder einer Combinationsreihe, in welcher die Entwicklungsreihen für je zwei Merkmale vereinigt sind.
Die vollkommene Uebereinstimmung, welche sämmtliche, dem Versuche unterzogenen Charaktere zeigen, erlaubt wohl und rechtfertigt die Annahme, dass auch ein gleiches Verhalten den übrigen Merkmalen zukomme, welche weniger scharf an den Pflanzen hervortreten und deshalb in die Einzelversuche nicht aufgenommen werden konnten. Ein Experiment über Blüthenstiele von verschiedener Länge gab im Ganzen ein ziemlich befriedigendes Resultat, obgleich die Unterscheidung und Einreihung der Formen nicht mit jener Sicherheit erfolgen konnte, welche für correcte Versuche unerlässlich ist.
Die Resultate, zu welchen die vorausgeschickten Versuche führten, veranlassten weitere Experimente, deren Erfolg geeignet erscheint, Aufschlüsse über die Beschaffenheit der Keim- und Pollenzellen der Hybriden zu geben. Einen wichtigen Anhaltspunkt bietet bei Pisum der Umstand, dass unter den Nachkommen der Hybriden constante Formen auftreten, und zwar in allen Combinirungen der verbundenen Merkmale. Soweit die Erfahrung reicht, finden wir es überall bestätigt, dass constante Nachkommen nur dann gebildet werden können, wenn die Keimzellen und der befruchtende Pollen gleichartig, somit beide mit der Anlage ausgerüstet sind, völlig gleiche Individuen zu beleben, wie das bei der normalen Befruchtung der reinen Arten der Fall ist. Wir müssen es daher als nothwendig erachten, dass auch bei der Erzeugung der constanten Formen an der Hybridpflanze vollkommen gleiche Factoren zusammenwirken. Da die vreschiedenen constanten Formen an einer Pflanze, ja in einer Blüthe derselben erzeugt werden, erscheint die Annahme folgerichtig, dass in den Fruchtknoten der Hybriden so vielerlei Keimzellen (Keimbläschen) und in den Antheren so vielerlei Pollenzellen gebildet werden, als constante Combinationsformen möglich sind, und dass diese Keim- und Pollenzellen ihrer inneren Beschaffenheit nach den einzelnen Formen entsprechen.
In der That lässt sich auf theoretischem Wege zeigen, dass diese Annahme vollständig ausreichen würde, um die Entwicklung der Hybriden in den einzelnen Generationen zu erklären, wenn man zugleich voraussetzen dürfte, dass die verschiedenen Arten von Keim- und Pollenzellen an der Hybride durchschnittlich in gleicher Anzahl gebildet werden.
Um diese Vorraussetzungen auf experimentellem Wege einer Prüfung zu unterziehen, wurden folgende Versuche ausgewählt: Zwei Formen, welche in der Gestalt der Samen und in der Färbung des Albumens constant verschieden waren, wurden durch Befruchtung verbunden.
Werden die differirenden Merkmale wieder mit A, B, a, b bezeichnet, so war:
| AB | Samenpflanze, | ab | Pollenpflanze, |
| A | Gestalt rund, | a | Gestalt kantig, |
| B | Albumen gelb, | b | Albumen grün. |
Die künstlich befruchteten Samen wurden sammt mehreren Samen der beiden Stammpflanzen angebaut, und davon die kräftigsten Exemplare für die wechselseitige Kreuzung bestimmt. Befruchtet wurde:
| 1. | die Hybride mit dem Pollen von | AB |
| 2. | die Hybride mit dem Pollen von | ab |
| 3. | AB mit dem Pollen der Hybriden | |
| 4. | ab mit dem Pollen der Hybriden |
Für jeden von diesen 4 Versuchen wurden an 3 Pflanzen sämmtliche Blüthen befruchtet. War die obige Annahme richtig, so mussten sich an den Hybriden Keim- und Pollenzellen von den Formen AB, Ab, aB, ab entwickeln und es wurden verbunden:
| 1. | die Keimzellen | AB, Ab, aB, ab | mit den Pollenzellen | AB |
| 2. | die Keimzellen | AB, Ab, aB, ab | mit den Pollenzellen | ab |
| 3. | die Keimzellen | AB | mit den Pollenzellen | AB, Ab, aB, ab |
| 4. | die Keimzellen | ab | mit den Pollenzellen | AB, Ab, aB, ab |
Aus jedem von diesen Versuchen konnten dann nur folgende Formen hervorgehen:
| 1. | AB, | ABb, | AaB, | AaBb |
| 2. | AaBb, | Aab, | aBb, | ab |
| 3. | AB, | ABb, | AaB, | AaBb |
| 4. | AaBb, | Aab, | aBb, | ab |
Wurden ferner die einzelnen Formen der Keim- und Pollenzellen von der Hybride durchschnittlich in gleicher Anzahl gebildet, so mussten bei jedem Versuche die angeführten vier Verbindungen in numerischer Beöziehung gleich stehen. Eine vollkommene Uebereinstimmung der Zahlenverhältnisse war indessen nicht zu erwarten, da bei jeder Befruchtung auch bei der normalen, einzelne Keimzellen unentwickelt bleiben oder später verkümern, und selbst manche von den gut ausgebildeten Samen nach dem Anbaue nicht zum Keimen gelangen. Auch beschränkt sich die gemachte Voraussetzung daruaf, dass bei der Bildung der verschiedenartigen Keim- und Pollezellen die gleiche Anzahl angestrebt werde, ohne dass diese an jeder einzelnen Hybride mit mathematischer Genauigkeit erreicht werden müsste.
Der erste und zweite Versuch hatten vorzugsweise den Zweck, die Beschaffenheit der hybriden Keimzellen zu prüfen, sowie der dritte und vierte Versuch über die Pollenzellen zu entscheiden hatte. Wie aus der obigen Zusammenstellung hervorgeht, mussten der erste und dritte Versuch, ebenso der zweite und vierte ganz gleiche Verbindungen liefern, auch sollte der Erfolg schon im zweiten Jahre an der Gestalt und Färbung der künstlich befruchteten Samen theilweise ersichtlich sein. Bei dem ersten und dritten Versuche kommen die dominirenden Merkmale der Gestalt und Farbe A und B in jeder Verbindung vor, und zwar zum Theile constant, zum Theile in hybrider Vereiniung mit den recessiven Charakteren a und b, weshalb sie sämmtlichen Samen ihre Eigenthümlichkeit aufprägen müssen. Alle Samen sollten daher, wenn die Voraussetzung eine richtige war, rund und gelb erscheinen. Bei dem zweiten und dritten Versuche hingegen ist eine Verbindung hybrid in Gestalt und Farbe, daher sind die Samen rund und gelb; eine andere ist hybrid in der Gestalt und constant in dem recessiven Merkmale der Farbe, daher die Samen und rund und grün; die dritte ist constant in dem recessiven Merkmale der Gestalt und hybrid in der Farbe, daher die Samen kantig und gelb; die vierte ist constant in beiden recessiven Merkmalen, daher die Samen kantig und grün. Bei diesen beiden Versuchen waren daher viererlei Samen zu erwarten, nämlich runde gelbe, runde grüne, kantige gelbe, kantige grüne.
Die Ernte entsprach den gestellten Anforderungen vollkommen.
Es wurden erhalten bei dem
An einem günstigen Erfolge war nun kaum mehr zu zweifeln, die nächste Generation musste die endgültige Entscheidung bringen. Von den angebauten Samen kamen im folgenden Jahre bei dem ersten Versuche 90, bei dem dritten 87 Pflanzen zur Fruchtbildung; von diesen brachten bei dem
| Versuche | |||
|---|---|---|---|
| 1. | 3. | ||
| 20 | 25 | runde gelbe Samen | AB |
| 23 | 19 | runde gelbe und grüne Samen | ABb |
| 25 | 22 | runde und kantige gelbe Samen | AaB |
| 22 | 21 | runde und kantige, gelbe und grüne Samen | AaBb |
| Bei dem zweiten und vierten Versuche gaben die runden und gelben Samen Pflanzen mit runden und kantigen, gelben und grünen Samen | AaBb |
| von den runden und grünen Samen wurden Pflanzen erhalten mit runden und kantigen grünen Samen | Aab |
| die kantigen gelben Samen gaben Pflanzen mit kantigen gelben und grünen Samen | aBb |
| aus den kantigen grünen Samen wurden Pflanzen gezogen, die wieder nur kantige grüne Samen brachten | ab |
Obwohl auch bei diesen beiden Versuchen einige Samen nicht keimten, konnte dadruch in den schon im vorhergehenden Jahre gefundenen Zahlen nichts geändert werden, da jede Samenart Pflanzen geb, ie in Bezug auf die Samen unter sich gleich und von den anderen verschieden waren. Es brachten daher:
| 2.Versuch | 4. Versuch | |||
|---|---|---|---|---|
| Pflanzen Samen von der Form | AaBb | |||
| Pflanzen Samen von der Form | Aab | |||
| Pflanzen Samen von der Form | aBb | |||
| Pflanzen Samen von der Form | ab | |||
Bei allen Versuchen erschienen daher sämmtliche Formen, welche die gemachte Voraussetzung verlangte, und zwar in nahezu gleicher Anzahl.
Bei einer weiteren Probe wurden die Merkmale der Blüthenfarbe und Axenlänge in die Versuche aufgenommen und die Auswahl so getroffen, dass im dritten Versuchsjahre jedes Merkmal an der Hälfte sämmtlicher Pflanzen hervortreten musste, falls die obige Annahme ihre Richtigkeit hatte. A, B, a, b dienen wieder zur Bezeichnung der verschiedenen Merkmale.
| A | Blüthen violett- roth, | a | Blüthen weiss, |
| B | Axe lang, | b | Axe kurz. |
Die Form Ab wurde befruchtet mit ab, woraus die Hybride Aab hervorging. Ferner wurde befruchtet aB gleichfalls mit ab, daraus die Hybride aBb. Im zweiten Jahre wurde für die weiteren Befruchtung die Hybride Aab als Samenpflanze, die andere aBb als Pollenpflanze verwendet.
| Samenpflanze | Aab, | Pollenpflanze | aBb, |
| mögliche Keimzellen | Ab, ab, | Pollenzellen | aB, ab. |
Aus der Befruchtung zwischen den möglichen Keim- und Pollenzellen mussten vier Verbindungen hervorgehen, nämlich:
Daraus wird ersichtlicht, dass nach obiger Vorraussetzung im dritten Versuchtsjahre von sämmtlichen Pflanzen
| die Hälfte violett-rothe Blüthen haben sollte (Aa) | Glieder : 1,3 |
| die Hälfte weisse Blüthe (a) | Glieder : 2,4 |
| die Hälfte eine lange Axe (Bb) | Glieder : 1,2 |
| die Hälfte eine kurze Axe (b) | Glieder : 3,4 |
Aus 45 Befruchtungen des zweiten Jahres wurden 187 Samen erhalten, wovon im dritten Jahre 166 Pflanzen zur Blüthe gelangten. Darunter erschienen die einzelnen Glieder in folgender Anzahl:
| Glied: | Blüthenfarbe: | Axe: | |
|---|---|---|---|
| violett-roth | lang | 47 Mal | |
| weiss | lang | 40 Mal | |
| violett-roth | kurz | 38 Mal | |
| weiss | kurz | 41 Mal |
Es kam daher
| die violett-rothe Blüthenfarbe | (Aa) | an 85 Pflanzen vor |
| die weisse Blüthenfarbe | (a) | an 81 Pflanzen vor |
| die lange Axe | (Bb) | an 87 Pflanzen vor |
| die kurze Axe | (b) | an 79 Pflanzen vor |
Die aufgestellte Ansicht findet auch in diesem Versuche eine ausreichende Bestätigung.
Für die Merkmale der Hülsenform, Hülsenfarbe und Blüthenstellung wurden ebenfalls Versuche im Kleinen angestellt und ganz gleich stimmende Resultate erhalten. Alle Verbindungen, welche durch die Vereinigung der verschiedenen Merkmale möglich wurden, erschienen pünktlich und in nahezu gleicher Anzahl.
Es ist daher auf experimentellem Wege die Annahme gerechtfertigt, dass die Erbsenhybriden Keim- und Pollenzellen bilden, welche ihre Beschaffenheit nach in gleicher Anzahl allen constanten Formen entsprechen, welche aus der Combinirung der durch Befruchtung vereinigten Merkmale hervorgehen.
Die Verschiedenheit der Formen unter den Nachkommen der Hybriden, sowie die Zahlenverhältnisse, in welchen dieselben beobachtet werden, finden in dem eben erwiesenen Satze eine hinreichende Erklärugn. Den einfachsten Fall bietet die Entwicklungsreihe für je zwei differirende Merkmale. Diese Reihe wird bekanntlich durch den Ausdruck: A+2Aa+a bezeichnet, wobei A und a die Formen mit den constant differirenden Merkmalen und Aa die Hybridgestalt beider bedeuten. Sie enthält unter drei verschiedenen Gliedern vier Individuen. Bei der Bildung derselben werden Pollen- und Keimzellen von der Form A und a durchschnittlich zu gleichen Theilen in die Befruchtung treten, daher jede Form zweimal, da vier Individuen gebildet werden. Es nehmen demnach an der Befruchtung theil:
| die Pollenzellen | A + A + a + a
| die Keimzellen | A + A + a + a
| |
Es bleibt ganz dem Zufalle überlassen, welche von den beiden Pollenarten sich mit jeder einzelnen Keimzelle verbindet. Indessen wird es nach den Regeln der Wahrscheinlichkeit im Durchschnitte vieler Fälle immer geschehen, dass sich jede Pollenform A und a gleich oft mit jeder Keimzellform A und a vereinigt; es wird daher eine von den beiden Pollenzellen A mit einer Keimzelle A, die andere mit einer Keimzelle a bei der Befruchtung zusammentreffen, und ebenso eine Pollenzelle a mit einer Keimzelle A, die andere mit a verbunden werden.
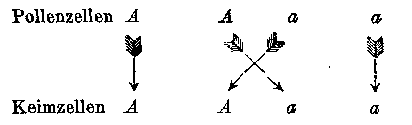
Das Ergebniss der Befruchtung lässt sich dadruch anschaulich machen, dass die Beziehungen für die verbundenen Keim- und Pollenzellen in Bruchform angesetzt werden, und zwar für die Pollenzellen über, für die Keimzellen unter dem Striche. Man erhält in dem vorliegenden Falle:
Bei dem ersten und vierten Gliede sind Keim- und Pollenzellen gleichartig, daher müssen die Producte ihrer Verbindung constant sei, nämlich A und a; bei dem zweiten und dritten hingegen erfolgt abermals eine Vereinigung der beiden differirenden Stamm-Merkmale, daher auch die aus diesen Befruchtungen hervorgehenden Formen mit der Hybride, von welcher sie abstammen, ganz identisch sind. Es findet demnach eine wiederholte Hybridisirung statt. Daraus erklärt sich die auffallende Erscheinung, dass die Hybriden im Stande sind, nebst den beiden Stammformen auch Nachkommen zu erzeugen, die ihnen gleich sind; und
geben beide dieselbe Verbindung Aa, da es, wie schon früher angeführt wurde, für den Erfolg der Befruchtung keinen Unterschied mach, welches von den beiden Merkmalen der Pollen- oder Keimzelle angehört. Es ist daher
So gestaltet sich der mittlere Verlauf bei der Selbstbefruchtung der Hybriden, wenn in denselben zwei differirende Merkmale vereinigt sind. In einzelnen Blüthen und an einzelnen Pflanzen kann jedoch das Verhältniss, in welchem die Formen der Reihe gebildet werden, nicht unbedeutende Störungen erleiden. Abgesehen davon, dass die Anzahl, in welcher beiderlei Keimzellen im Fruchtknogen vorkommen, nur im Durchschnitte als gleich angenommen werden kann, bleibt es ganz dem Zufalle überlassen, welche von den beiden Pollenarten an jeder einzelnen Keimzelle die Befruchtung vollzieht. Deshalb müssen die Einzelwerthe nothwendig Schwankungen unterliegen, und es sich selbst extreme Fälle möglich, wie sie früher bei den Versuchen über die Gestalt der Samen und die Färbung des Albumens angeführt wurden. Die wahren Verhältnisszahlen können nur durch das Mittel gegeben werden, welches aus der Summe möglichst vieler Einzelwerthe gezogen wird; je grösser ihre Anzahl, desto genauer wird das blos Zufällige eliminirt.
Die Entwicklungsreihe für Hybriden, in denen zweierlei differirende Merkmale verbunden sind, enthält unter 16 Individuen 9 verschiedene Formen, nämlich AB + Ab + aB + ab + 2ABb + 2aBb + 2AaB + 2Aab + 4AaBb. Zwischen den verschiedenen Merkmalen der Stammpflanzen A, a und B, b sind 4 constante Combinationen möglich, daher erzeugt auch die Hybride die entsprechenden 4 Formen von Keim- und Pollenzellen; AB, Ab, aB, ab und jede davon wird im Durchschnitte 4 Mal in Befruchtung treten, da in der Reihe 16 Individuen enthalten sind. Daher nehmen an der Befruchtung theil die
| Pollenzellen: | AB + AB + AB + AB + Ab + Ab + Ab + Ab + aB + aB + aB + aB + ab + ab + ab + ab |
| Keimzellen: | AB + AB + AB + AB + Ab + Ab + Ab + Ab + aB + aB + aB + aB + ab + ab + ab + ab |
Im mittleren Verlaufe der Befruchtung verbindet sich jede Pollenform gleich oft mit jeder Keimzellform, daher jede von den 4 Pollenzellen AB einmal mit einer von den Keimzellarten AB, Ab, aB, ab. Genau ebenso erfolgt die Vereinigung der übrigen Pollenzellen von den Formen Ab, aB, ab mit allen anderen Keimzellen. Man erhält demnach:
oder
In ganz ähnlicher Weise erklärt sich die Entwicklungsreihe der Hybriden, wenn in denselben dreierlei differirende Merkmale verbunden sind. Die Hybride bildet acht verschiedenen Formen von Keim- und Pollenzellen: ABC, ABc, AbC, Abc, aBC, aBc, abC, abc, und jede Pollenform vereinigt sich wieder durchschnittlich einmal mit jeder Keimzellform.
Das Gesetz der Combinirung der differirenden Merkmale, nach welchem die Entwicklung der Hybriden erfolgt, findet demnach seine Begründung und Erklärung in dem erwiesenen Satze, dass die Hybriden allen constanten Formen entsprechen, die aus der Combinirung der durch Befruchtung vereinigten Merkmale hervorgehen.
Es wird die Aufgabe weiterer Versuche sein, zu ermitteln, ob das für Pisum gefundene Entwicklungsgesetz auch bei den Hybriden anderer Pflanzen Geltung habe. Zu diesem Zwecke wurden in der letzten Zeit mehrere Versuche eingeleitet. Beendet sind zwei kleinere Experimente mit Phaseolusarten, welche hier Erwähnung finden mögen.
Ein Versuche mit Phaseolus vulgaris und Phaseolus nanus L. gab ein gaz übereinstimmendes Resultat. Ph. nanus hatte nebst der zwergartigen Axe grüne einfach gewölbte Hülsen, Ph. vulgaris hingegen ein 10-12' hohe Axe und gelb gefärbte, zur Zeit der Reife eingeschnürte Hülsen. Die Zahlenverhältnisse, in welchen die verschiedenen Formen in dem einzelnen Generationen vorkamen, waren dieselben wie bei Pisum. Auch die Entwicklung der constanten Verbindungen erfolgte nach dem Gesetze der einfachen Combinirung der Merkmale, genau so, wie es bei Pisum der Fall ist. Es wurden erhalten:
| Constante Verbindung: | Axe: | Farbe der unreifen Hülse: | Form der reifen Hülse: |
|---|---|---|---|
| gewölbt | |||
| eingeschnürt | |||
| gewölbt | |||
| eingeschnürt | |||
| gewölbt | |||
| eingeschnürt | |||
| gewölbt | |||
| eingeschnürt |
Die Grüne Hülsenfarbe, die gewölbte Form der Hülse und die hohe Axe waren, wie bei Pisum, dominirende Merkmale.
Ein anderer Versuch mit zwei sehr verschiedenen Phaseolusarten hatte nur einen theilweisen Erfolg. Als Samenpflanze diente Ph. nanus L., eine ganz constante Art mit weissen Blüthen in kurzen Trauben und kleinen weissen Samen in geraden, gewölbten und glatten Hülsen; als Pollenpflanze Ph. multiflorus W. mit hohem windenden Stengel, purpurrothen Blüthen in sehr langen Trauben, rauhen sichelförmig gekrümmten Hülsen und grossen Samen, welche auf pfirsichblüthrothem Grunde schwarz gefleckt und geflammt sind.
Die Hybride hatte mit der Pollenpflanze die grösste Aehnlichkeit, nur die Blüthen erschienen weniger intensiv gefärbt. Ihre Fruchtbarkeit war eine sehr beschränkte, von 17 Pflanzen, die zusammen viele hundert Blüthen entwickelten, wurden im Ganzen nur 49 Samen geerntet. Diese waren von mittlerer Grösse und besassen eine ähnliche Zeichnung wie Ph. multiflorus; auch die Grundfarbe war nicht wesentlich verschieden. Im nächsten Jahre wurden davon 44 Pflanzen erhalten, von denen nur 31 zur Blüthe gelangten. Die Merkmale von Ph. nanus, welche in der Hybride sämmtlich latent wurden, kamen in verschiedenen Combinirungen wieder zum Vorscheine, das Verhältniss derselben zu den dominirenden musste jedoch bei der geringen Anzahl von Versuchspflanzen sehr schwankend bleiben; bei einzelnen Merkmalen, wie bei jenen der Axe und der Hülsenform, war dasselbe indessen wie bei Pisum fast genau 1 : 3.
So gering der Erfolg dieses Versuches für die Feststellung der Zahlenverhältnisse sein mag, in welchen die verschiedenen Formen vorkamen, so bietet er doch andrerseits den Fall einer merkwürdigen Farbenwandlung an den Blüthen und Samen der Hybriden dar. Bei Pisum treten bekanntlich die Merkmale der Blüthen- und Samenfarbe in der ersten und den weiteren Generationen unverändert hervor und die Nachkommen der Hybriden tragen ausschliesslich das eine oder das anderen der beiden Stamm-Merkmale an sich. Anders verhält sich die Sache bei dem vorliegenden Versuche. Die weisse Blumen- und Samenfarbe von Ph. nanus erschien allerdings gleich in der ersten Generation an einem ziemlich fruchtbaren Exemplare, allein die übrigen 30 Pflanzen entwickelten Blüthenfarben, die verschiedenen Abstufungen von Purpurroth bis Blassviolett darstellen. Die Färbung der Samenschale war nicht minder verschieden, als die der Blüthe. Keine Pflanze konnte als vollkomen fruchtbar gelten, manche setzten gar keine Früchte an, bei anderen entwickelten sich dieselben erst aus den letzten Blüthen und kamen nicht mehr zur Reife, nur von 15 Pflanzen wurden gut ausgebildete Samen geerntet. Die meiste Neigung zur Unfruchtbarkeit zeigten die Formen mit vorherrschend rother Blüthe, indem von 16 Pflanezn nur 4 reife Samen gaben. Drie davon hatten ein ähnliche Samenzeichnung wie Ph. multiflorus, jedoch eine mehr oder weniger blasse Grundfarbe, die vierte Pflanze brachte nur einen Samen von einfach brauner Färbung. Die Formen mit überwiegend violetter Blüthenfarbe hatten dunkelbraune, schwarzbraune und ganz schwarze Samen.
Der Versuche wurde noch durch zwei Generationen unter gleich ungünstigen Verhältnissen fortgeführt, da selbst unter den Nachkommen ziemlich fruchtbarer Pflanzen wieder ein Theil wenig fruchtbar oder ganz steril wurde. Andere Blüthen- und Samenfarben, als die angeführten, kamen weiter nicht vor. Die Formen, welch ein der ersten Generation eines oder mherer von den recessiven Merkmalen erhielten, bleiben in Bezug auf diese ohne Ausnahme constant. Auch von jenen Samen Pflanzen, welche violette Blüthen und braune oder schwarze Samen besassen, änderten einzelne in den nächsten Generationen die Blumen- und Samenfarbe nicht mehr, die Mehrzahl jedoch erzeugte nebst ganz gleichen Nachkommen auch solche, welche weisse Blüthen und eben so gefärbte Samenschalen erhielten. Die rothblüthenden Pflanzen blieben so wenig fruchtbar, dass sich über ihre Weiterentwickelung nichts mit Bestimmtheit sagen lässt.
Ungeachtet der vielen Störungen, mit welchen die Beobachtung zu kämpfen hatte, geht doch so viel aus diesem Versuche hervor, dass die Entwicklung der Hybriden in Bezug auf jene Merkmale, welche die Gestalt der Pflanze betreffen, noch demselben Gesetze wie bei Pisum erfolgt. Rücksichlich der Farbenmerkmale scheint es allerdings schwierig zu sein, eine genügende Uebereinstimmung aufzufinden. Abgesehen davon, dass aus der Verbindung einer weissen und purpurrothen Färbung eine ganze Reihe von Farben hervorgeht, von Purpur bis Blassviolett und Weiss, muss auch der Umstand auffallen, dass unter 31 blühenden Pflanzen zur eine den recessiven Charakter der weissen Färbung erhielt, während das bei Pisum durchschnittlich schon bei jeder vierten Pflanze der Fall ist.
Auch auch diese räthsalhaften Erscheinungen würden sich
wahrscheinlich nach dem für Pisum geltenden Gesetze
erklären lassen, wenn man voraussetzen dürfte, dass die
Blumen- und Samenfarbe des Ph. multiflorus aus zwei oder mehreren
ganz selbstständigen Farben zusammengesetzt sei., die sich einzeln
ebenso verhalten, wie jedes andere constante Merkmal an der Pflanze.
Wäre die Blüthenfarbe A zusammengesetzt aus den
selbstständigen Merkmalen A1 +
A
Die Glieder dieser Reihen können in 9 verschiedene Verbindungen treten und jede davon stellt die Bezeichnung für eine andere Farbe vor:
| 1 A1A2 | 2 A1aA2 | 1 A2a |
| 2 A1A2a | 4 A1aA2a | 2 A2aa |
| 1 A1a | 2 A1aa | 1 aa |
Die den einzelnen Verbindungen vorausgesetzten Zahlen geben zugleich an, wie viele Pflanzen mit der entsprechenden Färbung in die Reihe gehören. Sa die Summe derselben 16 beträgt, so sind sämmtliche Farben im Durchschnitte auf je 16 Pflanzen vertheilt, jedoch, wie die Reihe selbst zeigt, in ungleichen Verhältnissen.
Würde die Farbenentwicklung wirklich in dieser Weise erfolgen, so könnte auch der oben angeführte Fall eine Erklärung finden, dass nämlich die weisse Blüthen- und Hülsenfärbe und 31 Pflanzen der ersten Generation nur einmal enthalten, und könnte daher auch nur im Durchschnitte unter je 16, bei drei Farbenmerkmalen sogar nur unter 64 Pflanzen einmal entwickelt werden.
Es darf jedoch nicht vergessen werden, dass die hier vorsuchte Erklärung auf einer blossen Vermuthung beruht, die weiter nichts für sich hat, als das sehr unvollständige Resultat des eben besprochenen Versuches. Es wäre übrigens eine lohnende Arbeit, die Farbenentwicklung der Hybriden durch ähnliche Versuche weiter zu verfolgen, da es wahrscheinlich ist, dass wir auf diesem Wege die ausserordendliche Mannigfaltigkeit in der Färbung unserer Zierblumen begreifen lernen.
Bis jetzt ist mit Sicherheit kaum mehr bekannt, als dass die Blüthenfarbe bei den meisten Zierpflanzen ein äusserst veränderliches Merkmal ist. Man hat häufig die Meinung ausgesprochen, dass die Stabilität derArten durch die Cultur in hohem Grade erschüttert oder ganz gebrochen werden, und ist sehr geneigt, die Entwicklung der Culturformen als eine regellose und zufällige hinzustellen; dabei wird gewöhnlich auf die Färbung der Zierpflanzen, als Muster aller Unbeständgkeit, hingewiesen. Es ist jedoch nicht einzusehen, warum das blosse Versetzen in den Gartengrund eine so durchgreifende und nachhaltige Revolution im Pflanzenorganismus zur Folge haben müsse. Niemand wird im erste behauptene wollen, dass die Entwicklung der Pfalnze im freien Lange durch andere Gesetze geleitet wird, als im Gartenbeete. Hier wie dort müssen typische Abänderungen auftreten, wenn die Lebensbedingungen für eine Art geändert werden und diese die Fähigkeit besitzt, sich den neuen Verhältnissen anzupasssen.Es wird gerne zugegeben, dass durch die Cultur die Entstehung neuer Varietäten begünstigt und durch die Hand des Menschen manche Abänderung erhalten wird, welche im freien Zustande unterliegen müsste, allein nichts berechtigt uns zu der Annahme, dass die Neigung zur Varietätenbildung so ausserordentlich gesteigert werde, dass die Arten bald alle Selbstständigkeit verlieren und ihre Nachkommen in einer endlosen Reihe höchst veränderlicher Formen aus einander gehen. Wäre die Aenderung in den Vegetationsbedingungen die alleinige Ursache der Variabilität, so dürfte man erwarten, dass jene Culturpflanzen, welche Jahrhunderte hindurch unter fast gleichen Verhältnissen angebaut wurden, wieder Selbstständigkeit gewonnen hätten. Das ist bekanntlich nicht der Fall, da gerade unter diesen nicht bloss die verschiedensten, sondern auch die veränderlichsten Formen gefunden werden. Nur die Leguminosen wie Pisum, Phaseolus, Lens, deren Befruchtungsorgane durch das Schiffchen geschützt sind, machen davon eine bemerkenswerthe Ausnahme. Auch da sind während einer mehr als 1000jährigen Cultur unter den mannigfaltigsten Verhältnissen zahlreiche Varietäten entstanden, diese behaupten jedoch unter gleich bleibenden Lebensbedingungen eine Selbstständigkeit, wie sie wild wachsenden Arten zukommt.
Es bleibt mehr als wahrscheinlich, dass für die Veränderlichkeit der Culturgewächse ein Factor thätig ist, dem bisher wenig Aufmerksamkeit zugewendet wurde. Verschiedene Erfahrungen drängen zu der Ansicht, dass unsere Culturpflanzen mit wenigen Ausnahmen Glieder verschiedener Hybridreihen sind, deren gesetzmäßige Weiterentwicklung durch häufige Zwischenkreuzungen abgeändert und aufgehalten wird. Es ist der Umstand nicht zu übersehen, dass die cultivirten Gewächse meistens in grösserer Anzahl neben einander gezogen werden, wodruch für die wechselseitige Befruchtung zwischen vorhandenen Varietäten und mit den Arten selbst die günstigste Gelegenheit geboten wird. Die Wahrscheinlichkeit dieser Ansicht wird durch die Thatsache unterstützt, dass unter dem grossen heere veränderlicher Formen immer einzelne gefunden werden, welche in dem einen oder anderen Merkmale constant bleiben, wenn nur jeder fremde Einfluss sorgfältig abgehalten wird. Diese Formen entwickeln sich genau eben so, wie gewisse Glieder der zusammengesetzten Hybridreihen. Auch bei dem empfindlichsten aller Merkmale, bei jenem der Farbe, kann es der aufmerksamen Beobachtung nicht entgehen, dass an den einzelnen Formen die Neigung zur Veränderlichkeit in sehr verschiedenem Grade vorkommt. Unter Pflanzen, die aus einer spontanen Befruchtung stammen, giebt es oft solche, deren Nachkommen in Beschaffenheit und Anordnung der Farben weit auseinandergehen, während andere wenig abweichende Formen liefern, und unter einer grösseren Anzahl einzelne getroffen werden, welche ihre Blumenfarbe unverändert auf die Nachkommen übertragen. Die cultivirten Dianthusarten geben dafür einen lehrreichen Beleg. Ein weiss blühendes Exemplar von Dianthus Caryophyllus, welches selbst von einer weissblühenden Varietät abstammte, wurde während der Blüthezeit in einem Glasshause eingesperrt; die zahlreich davon gewonnenen Samen gaben Pflanzen mit durchaus gleicher weisser Blüthenfarbe. Ein ähnliches Resultat wurde von einer rothen, etwas ins Violette schimmernden und einer weissen, roth gestreiften Abart erhalten. Viele andere hingegen, welche auf dieselbe Weise geschützt wurden, gaben mehr oder weniger verschieden gefärbte und gezeichnete Nachkommen.
Wer die Färbungen, welche bei Zierpflanzen aus gleicher Befruchtung hervorgehen, überblickt, wird sich nicht leicht der Ueberzeugung verschliessen können, dass auch hier die Entwicklung nach einem bestimmten Gesetze erfolgt, welches möglicherweise seinen Ausdruck in der Combinirung mehrerer selbstständiger Farbenmerkmale findet.
Es dürfte nicht ohne Interesse sein, die bei Pisum gemachten Beobachtungen mit den Resultaten zu vergleichen, zu welchen die beiden Autoritäten in diesem Fache, Kölreuther und Gärtner, bei ihren Forschungen gelangt sind. Nach der übereinstimmenden Ansicht beider halten die Hybriden der äusseren Erscheinung nach entweder die Mittelform zwischen den Stammarten, oder sie sind dem Typus der einen oder der anderen näher gerückt, manchmal von denselben kaum zu unterscheiden. Aus den Samen derselben gehen gewöhnlich, wenn die Befruchtung durch den eigenen Pollen geschah, verschiedene von dem normalen Typus abweichende Formen hervor. In der Regel behält die Mehrzahl der Individuen aus einer Befruchtung die Form der Hybride bei, während andere wenige der Samenpflanze ähnlicher werden und ein oder das andere Individuum der Pollenpflanze nahe kommt. Das gilt jedoch nicht von allen Hybriden ohne Ausnahme. Bei einzelnen sind die Nachkommen theils der einen theils der anderen Stammpflanze nähre gerückt, oder sie neigen sich sämmtlich mehr nach der einen oder der anderen Seite hin; bei einigen aber bleiben sie der Hybride vollkommen gleich und pflanzen sich unverändert fort. Die Hybriden der Varietäten verhalten sich wie die Specieshybriden, nur besitzen sie eine noch grössere Veränderlichkeit der Gestalten und eine mehr ausgesprochene Neigung, zu den Stammformen zurück zu kehren.
In Bezug auf die Gestalt der Hybriden und ihre in der Regel erfolgende Entwicklung ist eine Uebereinstimmung mit den bei Pisum gemachten Beobachtungen nicht zu verkennen. Anders verhält es sich mit den erwähnten Ausnahmsfällen. Gärtner gesteht selbst, dass die genaue Bestimmung, ob eine Form mehr der einen oder der anderen von den beiden Stammarten ähnlich sei, öfter grosse Schwierigkeiten habe, indem dabei sehr viel auf die subjective Anschauung des Beobachters ankommt. Es konnte jedoch auch ein anderer Umstand dazu beibetragen, dass die Resultate trotz der sorgfältigsten Beobachtung und Unterscheidung schwankend und unsicher wurden. Für die Versuche dienten grösstentheils Pflanzen, welche als gute Arten gelten und in einer grösseren Anzahl von Merkmalen verschieden sind. Nebst den scharf hervortretenden Charakteren müssen da, wo es sich im Allgemeinen um eine grössere oder geringere Aehnlichkeit handelt, auch jene Merkmale eingerechnet werden, welche oft schwer mit Worten zu fassen sind, aber dennoch hinreichen, wie jeder Pflanzenkenner weiss, um den Formen ein fremdartiges Aussehen zu geben. Wird angenommen, dass die Entwicklung der Hybriden nach dem für Pisum geltenden Gesetze erfolgte, so musste die Reihe bei jedem einzelnen Versuche sehr viele Formen umfassen, da die Gliederzahl bekanntlich mit der Anzahl der differirenden Merkmale nach den Potenzen von 3 zunimmt. Bei einer verhältnissmässig kleinen Anzahl von Versuchspflanzen konnte dann das Resultat nur annähernd richtig sein und in einzelnen Fällen nicht unbedeutend abweichen. Wären z. B. die beiden Stammarten in 7 Merkmalen verschieden und würden aus den Samen ihrer Hybriden zur Beurtheilung des Verwandtschaftsgrades der Nachkommen 100 bis 200 Pflanzen gezoen, so sehen wir leicht ein, wie unsicher das Urtheil ausfallen müsste, da für 7 differirende Merkmale die Entwicklungsreihe 16,384 Individuen unter 2187 verschiedenen Formen enthält. Es könnte sich bald die eine, bald die andere Verwandtschaft mehr geltend machen, je nchdem der Zufall dem Beobachter diese oder jene Formen in grösserer Anzahl in die Hand spielt.
Kommen ferner unter den differirenden Merkmalen zugleich dominirende vor, welche ganz oder fast unverändert auf die Hybride übergehen, dann muss an den Glieder der Entwicklungsreihe imre jene der beiden Stammarten mehr hervortreten, welche die grössere Anzahl der dominirenden Merkmale besitzt. In dem früher bei Pisum für dreierlei differirende Merkmale angeführten Versuche gehörten die dominirenden Charaktere sämmtlich der Samenpflanze an. Obwohl die Glieder der Reihe sich ihrer inneren Beschaffenheit nach gleichmässig zu beiden Stammpflanzen hinneigen, erhielt doch bei diesem Versuche der Typus der Samenpflanze ein so bedeutendes Uebergewicht, dass unter je 64 Pflanzen der ersten Generation 54 derselben ganz gleich waren. Man sieht, wie gewagt es unter Umständen sein kann, bei Hybriden aus der äusseren Uebereinstimmung Schlüsse auf ihre innere Verwandtschaft zu ziehen.
Gärtner erwähnt, dass in jenen Fällen, wo die Entwicklung eine regelmässgie war, unter den Nachkommen der Hybriden nicht die beiden Stammarten selbst erhalten wurden, sondern nur einzelne ihnen näher verwandte Individuen. Bei nicht sehr ausgedehnten Entwicklungsreihen konnte es in der That nicht anders eintreffen. Für 7 differirende Merkmale z. B. kommen unter mehr als 16,000 Nachkommen der Hybride die beiden Stammformen nur je einmal vor. Es ist demnach nicht leicht möglich, dass dieselben schon unter einer geringen Anzahl von Versuchspflanzen erhalten werden; mit einiger Wahrscheinlichkeit darf man jedoch auf das Erscheinen einzelner Formen rechnen, die demselben in der Reihe nahe stehen.
Einer wesentlichen Verschiedenheit begegnen wir bei jenen Hybriden, welche in ihren Nachkommen constant bleiben und sich eben so wie die reinen Arten fortpflanzen. Nach Gärtner gehören hieher die ausgezeichnet fruchtbaren Hybriden: Aquilegia atropurpurea-canadensis, Lavatera speudolbia-thuringiaca, Geum urbano-rivale und einige Dianthushybriden; nach Wichura die Hybriden der Weidenarten. Für die Entwicklungsgeschichte der Pflanzen ist dieser Umstand von besonderer Wichtigkeit, weil constante Hybriden die Bedeutung neuer Arten erlangen. Die Richtigkeit des Sachverhaltes ist durch vorzügliche Beobachter verbürgt und kann nicht in Zweifel gezogen werden. Gärtner hatte Gelegenheit, den Dianthus Armeria-deltoides bis in die 10. Generation selbst zu verfolgen, da sich derselbe regelmässig im Garten von selbst fortpflanzte.
Bei Pisum wurde es durch Versuche erwiesen, dass die Hybriden verschiedenartige Keim- und Pollenzellen bilden, und dass hierin der Grund für die Veränderlichkeit ihrer Nachkommen liegt. Auch bei anderen Hybriden, deren Nachkommen sich ähnlich verhalten, dürfen wir eine gleiche Ursache voraussetzen; für jene hingegen, welche constant beiben, scheint die Annahme zulässig, dass ihre Befruchtungszellen gleichartig sind und mit der Hybriden-Grundzelle übereinstimmen. Nach der Ansicht berühmter Phyiologen vereinigen sich bei Phanerogamen zu dem Zwecke der Fortpflanzung je eine Keim- und Pollenzelle zu einer einzigen Zelle {Fussnote: Bei Pisum ist es wohl ausser Zweifel gestellt, dass zur Bildung des neuen Embryo eine vollständige Vereinigung der Elemente beider Befruchtungszellen stattfinden müsse. Wie wollte man es sonst erklären, dass unter den Nachkommen der Hybriden beide Stammformen in gleicher Anzahl und mit allen ihren Eigenthümlichkeiten wieder hervortreten? Wäre der Einfluss des Keimsackes auf die Pollenzelle nur ein äusserer, wäre demselben bloss die Rosse einer Amme zugetheilt, dann könnte der Erfolg einer jeden künstlichen Befruchtung kein anderer sein, als dass die entwickelte Hybride ausschliesslich der Pollenpflanze gleich käme, oder ihr doch sehr nahe stände. Das haben die bisherigen Versuche in keinerlei Weise bestätigt. Ein gründlicher Beweis für die vollkommene Vereinigung des Inhaltes beider Zellen liegt wohl in der allseitig bestätigten Erfahrung, dass es für die Gestalt der Hybride gleichgültig ist, welche von den Stammformen die Samen- oder Pollenpflanze war.}, welche sich durch Stoffaufnahme und Bildung neuer Zellen zu einem selbstständigen Organismus weiter zu entwickeln vermag. Diese Entwicklung erfolgt nach einem constanten Gesetze, welches in der materiellen Beschaffenheit und Anordnung der Elemente begründet ist, die in der Zelle zur lebensfähigen Vereinigung gelangten. Sind die Fortpflanzungszellen gleichartig und stimmen dieselben mit der Grundzelle der Mutterpflanze überein, dann wird die Entwicklung des neuen Individuums durch dasselbe Gesetz geleitet, welches für die Mutterpflanze gilt. Gelingt es, eine Keimzelle mit einer ungleichartigen Pollenzelle zu verbinden, so müssen wir annehmen, dass zwischen jenen Elementen beider Zellen, welche die gegenseitigen Unterschiede bedingen, irgend eine Ausgleichung stattfindet. Die daraus hervorgehende Vermittlungszelle wird zur Grundlage des Hybriden-Organismus, dessen Entwicklung nothwendig nach einem anderen Gesetze erfolgt, als bei jeder der beiden Stammarten. Wird die Ausgleichung als eine vollständige angenommen, i dem Sinne nämlich, dass der hybride Embryo aus gleichartigen Zellen gebildet wird, in welchen die Differenzen gänzlich und bleibend vermittelt sind, so würde sich als weitere Folgerung ergeben, dass die Hybride, wie jede andere selbstständige Pflanzenart, in ihren Nachkommen constant bleiben werde. Die Fortpflanzungszellen, welche in dem Fruchtknoten und den Antheren derselben gebildet werden, sind gleichartig und stimmen mit der zu Grunde liegenden Vermittlungszelle überein.
Bezüglich jener Hybriden, deren Nachkommen veränderlich sind, dürfte man vielleicht annehmen, dass zwischen den differirenden Elementen der Keim- und Pollenzelle wohl insofern eine Vermittlung stattfindet, dass noch die Bildung einer Zelle als Grundlage der Hybride möglich wird, dass jedoch die Ausgleichung der widerstrebenden Elemente nur eine vorübergehende sei und nicht über der Leben der Hybridpflanze hinausreiche. Da in dem Habitus derselben während der ganzen Vegetationsdauer keine Aenderungen wahrnehmbar sind, müssten wir weiter folgern, dass es den differirenden Elementen erst bei der Entwicklung der Befruchtungszellen gelinge, aus der erzwungenen Verbindung herauszutreten. Bei der Bildung dieser Zellen betheiligen sich alle vorhandenen Elemente in völlig freier und gleichmässiger Anordnung, wobei nur die differirenden sich gegenseitig ausschliessen. Auf diese Weise würde die Entstehung so vielerlei Keim- und Pollenzellen ermöglicht, als die bildungsfähigen Elemente Combinationen zulassen.
Die her versuchte Zurückführung des wesentlichen Unterschiedes in der Entwicklung der Hybriden auf eine dauernde oder vorübergehende Verbindung der differirenden Zellelemente kann selbstverständlich nur den Werth einer Hypothese ansprechen, für welche bei dem mangel an sicheren Daten noch ein weiterer Spielraum offen stände. Einige Berechtigung für die ausgesprochene Ansicht liegt in dem für Pisum angeführten Beweise, dass das Verhalten je zweier differirender Merkmale in hybrider Vereinigung unabhängig ist von den anderweitigen Unterschieden zwischen den beiden Stammpflanzen, und ferner, dass die Hybride so vielerlei Keim- und Pollenzellen erzeugt, als constante Combinationsformen möglich sind. Die unterscheidenden Merkmale zweier Pflanzen können zuletzt doch nur auf Differenzen in der Beschaffenheit und Gruppirung der Elemente beruhen, welche in den Grundzellen derselben in lebendiger Wechselwirkung stehen.
Die Geltung der für Pisum aufgestellten Sätze bedarf selbst noch der Bestätigung, und es wäre deshalb eine Wiederholung wenigstens der wichtigeren Versuche wünschenswerth, z. b. jener über die Beschaffenheit der hybriden Befruchtungszellen. Dem einzelnen Beobachter kann leicht ein Differentiale entgehen, welches, wenn es auch anfangs unbedeutend scheint, doch so anwachsen kann, dass es für das Gesammt- Resultat nicht vernachlässig werden darf. Ob die veränderlichen Hybriden anderer Pflanzenarten ein ganz übereinstimmendes Verhalten beobachten, muss gleichfalls erst durch Versuche entschieden werden; indessen dürfte man vermuthen, dass in wichtigen Punkten eine principielle Verschiedenheit nicht vorkommen könne, da die Einheit im Entwicklungsplane des organischen Lebens ausser Frage steht.
Zum Schlusse verdienen noch eine besondere Erwähnung die von Kölreuther, Gärtner u. a. durchgeführten Versuche über die Umwandlung einer Art in eine andere durch künstliche Befruchtung. Diesen Experimenten wurde eine besondere Wichtigkeit beigelegt, Gärtner rechnet dieselben zu den "allerschwierigsten in der Bastarderzeugung."
Sollte eine Art A in eine andere B verwandelt werden, so wurden beide durch Befruchtung verbunden und die erhaltenen Hybriden abermals mit dem Pollen von B befruchtet; dann wurd aus den verschiedenen Abkömmlingen derselben jene Form ausgewählt, welche der Art B am nächsten stand, und wiederholt mit dieser befruchtet, und so fort, bis man endlich eine Form erhielt, welche der B gleichkam und in ihren Nachkommen constant blieb. Damit war die Art A in die andere Art B umgewandelt. Gärtner hat allein 30 derartige Versuche mit Pflanzen aus den Geschlechtern: Aquilegia, Dianthus, Geum, Lavatera, Lychnis, Malva, Nicotiana und Oenothera durchgeführt. Die Umwandlungsdauer war nicht für alle Arten die gleiche. Während bei einzelnen eine 3malige Befruchtung hinreichte, musste diese bei einzelnen 5-6mal wiederholt werden; auch für die nämlichen Arten wurden bei verschiedenen Versuchen Schwankungen beobachtet. Gärtner schreibt diese Verschiedenheit dem Umstande zu, dass "die typische Kraft, womit eine Art bei der Zeugung zur Veränderung und Umbildung des mütterlichen Typus wirkt, bei den verschiedenen Gewächsen sehr verschieden sein müssen, und die Umwandlung bei manchen Arten durch mehr, bei anderen aber durch weniger Generationen vollbracht wird. Ferner bemerkt derselbe Beobachter, "dass es auch bei Umwandlungsgeschäfte darauf ankommt, welcher Typus und welches Individuum zu der weiteren Umwandlung gewählt wird."
Dürfte man voraussetzen, dass bei diesen Versuchen die Entwicklung der Formen auf eine ähnliche Weise wie bei Pisum erfolgte, so würde der ganze Umwandlungsprocess eine ziemlich einfache Erklärung finden. Die Hybride bildet so vielerlei Keimzellen, als die in ihr vereinigten Merkmale constante Combinationen zulassen, und eine davon ist immer gleichartig mit den befruchtenden Pollenzellen. Demnach ist für alle derartigen Versuche die Möglichkeit vorhanden, dass schon aus der zweiten Befruchtung eine constante Form gewonnen wird, welche der Pollenpflanze gleich kommt. Ob dieselbe aber wirklich erhalten wird, hängt in jedem einzelnen Falle von der Zahl der Versuchspflanzen ab, sowie von der Anzahl der differirenden Merkmale, welche durch die Befruchtung vereinigt wurden. Nehmen wir z. B. an, die für den Versuch bestimmten Pflanzen wären in 3 Merkmalen verschieden und es sollte die Art ABC in die andere abc durch wiederholte Befruchtung mit dem Pollen derselben umgewandelt werden. Die aus der ersten Befruchtung hervorgehende Hybride bildet 8 verschiedene Arten von Keimzellen, nämlich:
Diese werden im zweiten Versuchsjahre abermals mit dem Pollen abc verbunden und man erhält die Reihe:
Da die Form abc in der 8 gliedrigen Reihe einmal vorkommt, so ist es wenig wahrscheinlich, dass sie unter den Versuchspflanzen fehlen könnte, wenn diese auch nur in einer geringen Anzahl gezogen würden, und die Umwandlung wäre schon nach zweimaliger Befruchtung vollendet. Sollte sie zufällig nicht erhalten werden, so müsste die Befruchtung an einer der nächstverwandten Verbindungen Aabc, aBCc, abCc wiederholt werden. Es wird ersichtlich, dass sich ein derartiges Experiment desto länger hinausziehen müsse, je kleiner die Anzahl der Versuchspflanzen und je grösser die Zahl der differirenden Merkmale an den beiden Stammarten ist, dass ferner bei den nämlichen Arten leicht eine Verschiebung um eine, selbst um zwei Generationen vorkommen könne, wie es Gärtner beobachtet hat. Die Umwandlung weit abstehender Arten kann immerhin erst im 5. oder 6. Versuchsjahre beendet sein, indem die Anzahl der verschiedenen Keimzellen, welche an der Hybride gebildet werden, mit den differirenden Merkmalen nach den Potenzen von 2 zunimmt.
Gärtner fand durch wiederholte Versuche, dass die wechselseitige Umwandlungsdauer für manche Arten verschieden ist, so dass öfter eine Art A in eine andere B um eine Generation früher verwandelt werden kann, als die Art B in die andere A. Er leitet daraus zu den Beweis ab, dass die Ansicht Kölreuther's doch nicht ganz stichhaltig sei, nach welcher "die beiden Naturen bei den Bastarden einander das vollkommenste Gleichgewicht halten." Es scheint jedoch, dass Kölreuther diesen Tadel nicht verdient, dass vielmehr Gärtner dabei ein wichtiges Moment übersehen hat, auf welches er an einer anderen Stelle selbst aufmerksam mach, dass es nämlich darauf ankommt, welches Individuum zur weiteren Umwandlung gewählt wird". Versuche, welche in dieser Beziehung mit zwei Pisum-Arten angestellt wurden, weisen darauf hin, dass es für die Auswahl der tauglichsten Individuen zu dem Zwecke der weiteren Befruchtung einen grossen Unterschied machen könne, welche von zwei Arten in die andere umgewandelt wird. Die beiden Versuchspflanzen waren in 5 Merkmalen verschieden, zugleich besass die Art A sämmtliche dominirende, die andere B sämmtliche recessive Merkmale. Für die wechselseitige Umwandlung von A mit dem Pollen von B und umgekehrt B mit jenem von A befruchtet, dann dasselbe an beiderlei Hybriden im nächsten Jahre wiederholt. Bei dem ersten Versuche waren im 3. Versuchsjahre für die Auswahl der Individuen zur weiteren Befruchtung 87 Pflanzen vorhanden, und zwar in dem möglichen 32 Formen; für den zweiten Versuch wurden 73 Pflanzen erhalten, welche in ihrem Habitus durchgehends mit der Pollenpflanze übereinstimmten, jedoch ihrer inneren Beschaffenheit nach eben so verschieden sein mussten, wie die Formen des anderen Versuches. Eine berechnete Auswahl war daher bloss bei dem ersten Versuche möglich, bei dem zweiten mussten auf den blossen Zufall hin einige Pflanzen ausgeschieden werden. Von den letzteren wurde nur ein Theil der Blüthen mit den Pollen von A befruchtet, der andere hingegen der Selbstbefruchtung überlassen. Unter je 5 Pflanzen, welche für die beiben Versuche zur Befruchtung verwendet waren, stimmten, wie der nächstjahrige Anbau zeigte, mit der Pollenpflanze überein:
| Erster Versuch | Zweiter Versuch | |
|---|---|---|
| 2 Pflanzen | - | in allen Merkmalen |
| 3 Pflanzen | - | in 4 Merkmalen |
| - | 2 Pflanzen | in 3 Merkmalen |
| - | 2 Pflanzen | in 2 Merkmalen |
| - | 1 Pflanze | in 1 Merkmal |
Für den ersten Versuch war damit die Umwandlung beendet, bei dem zweiten, der nicht weiter forgesetzt wurde, hätte wahrscheinlich noch eine zweimalige Befruchtung stattfinden müssen.
Wenn auch der Fall nicht häufig vorkommen dürfte, dass die dominanten Merkmale ausschliesslich der einen oder der anderen Stammpflanze angehören, so wird es doch immer einen Unterschied machen, welche von beiden die grössere Anzahl besitzt. Kommt die Mehrzahl der dominirenden Merkmale der Pollenpflanze zu, dann wird die Auswahl der Formen für die weitere Befruchtung einen geringeren Grad von Sicherhiet gewähren, als in dem umgekehrten Falle, was eine Verzögerung in der Umwandlungsdauer zur Folge haben muss, vorausgesetzt, dass man den Versuch erst dann als beendet ansieht, wenn eine Form erhalten wird, die nicht nur in ihrer Gestalt der Pollenpflanze gleich kommt, sondern auch wie diese in den Nachkommen constant bleibt.
Durch den Erfolg der Umwandlungsversuche wurde Gärtner bewogen, sich gegen die Meinung derjenigen Naturforscher zu kehren, welche die Stabilität der Pflanzenspecies bestreiten und eine stete Fortbildung der Gewächsarten annehmen. Er sieht in der vollendeten Umwandlung einer Art in die andere den unzweideutigen Beweis, dass der Species feste Grenzen gesteckt sind, über welche hinaus sie sich nicht ändern vermag. Wenn auch dieser Ansicht eine bedingungslose Geltung nicht zuerkannt werden kann, so findet sich doch anderseits in den von Gärnter angestellten Versuchen eine beachtenswerthe Bestätigung der früher über die Veränderlichkeit der Culturpflanzen ausgesprochenen Vermuthung.
Unter den Versuchsarten kommen cultivirte Gewächse vor, wie Aquilegia atropupurea und canadensis, Dianthus Caryophyllus, chinensis und japonicus, Nicotiana rustica und paniculata, und auch diese hatten nach einer 4-5maligen Verbindung nichts von ihrer Selbstständigkeit verloren.
von
Gregor Mendel
(Mitgetheilt in der Sitzung vom 9. Juni 1869.)
Gedruckt in den Verhandlungen des naturforschenden Vereines in Brünn. VIII. Bd. Abhandlungen. 1869. Brünn, 1870. Verlag des Vereins. S. 26-31.
Wiewohl ich schon mehrfach Befruchtungsversuche zwischen verschiedenen Arten aus dem Genus Hieracium vorgenommen habe, ist es mir bis jetzt doch nur gelungen, folgende 6 Bastarde und diese bloss in einem bis drei Exemplaren zu erhalten:
| H. Auricula | + | H. aurantiacum *), |
| H. Auricula | + | H. Pilosella, |
| H. Auricula | + | H. pratense, |
| H. echoides**) | + | H. aurantiacum, |
| H. praealtum | + | H. flagellare Rchb., |
| H. praealtum | + | H. aurantiacum. |
{Fussnote: *) Durch diese Bezeichnung wird angedeutet, dass der Bastard aus der Befruchtung des H. Auricula mit dem Pollen des H. aurantiacum erhalten wurde.}
{Fussnote: **) Diese Versuchspflanze ist nicht genau das typische H. echioides. Sie scheint der Uebergangsreihe zu H. praealtum anzugehören, steht jedoch dem H. echioides näher, weshalb sie auch in den Formenkreis der letzteren eingestellt wurde.}
Die Schwierigkeit, Bastarde in einer grösseren Anzahl zu gewinnen, liegt in dem Umstande, dass es bei der Kleinheit der Blüthen und dem eigenthümlichen Baue derselben nur selten gelingt, die Antheren aus der zu befruchtenden Blüthe zu entfernen, ohne dass der eigene Pollen auf die Narbe gelangt, oder Griffel verletzt wird und abstirbt. Bekanntlich sind die Antheren in ein Röhrchen verwachsen, welches den Griffel eng umschliesst. Sobald die Blüthe sich öffnet, tritt die Narbe schon mit Pollen überdeckt aus dem Röhrchen hervor. Um die Selbstbefruchtung zu verhüten, muss deshalb das Antherenröhrchen noch vor dem Aufblühen entfernt und zu diesem Zwecke die Knospe mittelst einer feinen Nadel aufgeschlitzt werden. Wird diese Operation zu einer Zeit vorgenommen, wo der Pollen schon Befruchtungsfähigkeit erlangt hat, was 2-3 Tage vor dem Aufblühen der Fall ist, so gelingt es nur selten, die Selbstbefruchtung zu hindern, da es bei aller Aufmerksamkeit nicht leicht möglich ist, zu verhüten, dass bei dem Auschlitzen des Röhrchens einzelne Pollenkörner ausgestreut und der Narbe mitgetheilt werden. Keinen besseren Erfolg gewährte bis jetzt die Entfernung der Antheren in einem früheren Entwicklungstadium. Vor dem Eintritte der Pollenreife sind nämlich die noch sehr zarten Griffel und Narbe gegen Druck und Verletzungen äusserst empfindlich, und wenn sie auch nicht beschädigt wurden, welken und trocknen sie doch gewöhnlich nach kurzer Zeit ab, sobald sie ihrer schützenden Hüllen beraubt sind. Dem letzteren Uebelstande hoffe ich dadruch abzuhelfen, dass die Pflanze nach der Operation durch 2 bis 3 Tage der feuchten Atmosphäre des Warmhauses ausgesetzt wird. Ein Versuch, der vor Kurzem mit H. auricula in dieser Weise angestellt wurde, lieferte es gutes Resultat.
Um den Zweck anzudeuten, zu welchem die Befruchtungsversuche unternommen wurden, erlaube ich mir einige Bemerkungen über das Genus Hieracium vorauszuschicken. Dieses Genus besitzt einen so ausserordentlichen Reichthum an selbstständigen Formen, wie ihn kein anderes Pflanzengeschlecht aufweisen kann. Einzelne davon sind durch besondere Eigenthümlichkeiten ausgezeichnet und werden als Hauptformen oder Arten betrachtet, während alle übrigen sich als Mittelbildungen oder Uebergangsformen darstellen, durch welche die Hauptformen mit einander zusammenhängen. Die Schwierigkeit in der Gliederung und Abgrenzung dieser Formen hat die Aufmerksamkeit der Fachgelehrten immer in Anspruch genommen. Ueber keine anderen Gattung ist so viel geschrieben, sind so viele und heftige Kämpfe geführt worden, ohne dass es bis jetzt zu einem Abschlusse gekommen wäre. Es ist voraus zu sehen, dass einen Verständigung nicht zu erzielen sein wird, so lange nicht der Werth und die Bedeutung der Zwischen- oder Uebergangsformen erkannt ist.
Bezüglich der Frage, ob und in welchem Umfange die Bastardbildung an dem Formenreichthum des genannten Geschlechtes Antheil nimmt, begegnen wird unter den ersten Pflanzenkennern sehr abweichenden, sogar völlig widersprechenden Ansichten. Während einige derselben einen weit reichenden Einfluss zugestehen, wollen andere, z. B. Fries, bei Hieracien von Bastarden überhaupt nichts wissen. Noch andere nehmen eine vermittelnde Stellung ein und geben zu, dass Bastarde unter den wildwachsenden Arten nicht selten gebildet werden, behaupten jedoch, dass denselben eine wichtigere Bedeutung aus dem Grunde nicht beizumessen sei, weil sie immer nur von kurzem Bestande sind. Die Ursache davon liege theils in der geringen Fruchtbarkeit oder gänzlichen Sterilität derselben, theils aber in der durch Versuche erwiesenen Erfahrung, dass bei Bastarden die Selbstbefruchtung immer ausgeschlossen werde, wenn der Polln der Stammarten auf die Narben derselben gelangt. Es sei demnach undenkbar, dass Hieraciumbastarde sich in der Nähe ihrer Stammeltern zu vollkommen fruchtbaren und constanten Formen herausbilden und behaupten könnten.
Die Frage über den Ursprung der zahlreichen constanten Zwischenformen hat in neuester Zeit nicht wenig an Interesse gewonnen, seitdem ein berühmter Hieracienkenner im Geiste der Darwin'schen Lehre die Ansicht vertritt, dass dieselben aus der Transmutation untergegangener oder noch bestehender Arten herzuleiten seien.
Es liegt in der Sache, um die es sich hier handelt, dass eine genaue Kenntnis der Bastarde in Bezug auf ihre Gestalt und Fruchtbarkeit, sowie auf das Verhalten ihrer Nachkommen durch mehrere Generationen unerlässlich ist, wenn man es unternehmen will, den Einfluss zu beurtheilen, den möglicherweise die Bastardbildung auf die Mannigfaltigkeit der Zwischenformen bei Hieracium ausübt. Das Verhalten der Hieracium-Bastarde in dem angedeuteten Umfange muss nothwendig durch Versuche ermittelt werden, da wir eine abgeschlossenen Theorie der Bastardbildung nicht besitzen, und es zu irrigen Anschauungen führen könnte, wenn man die aus der Beobachtung einiger anderer Bastarde abgeleiteten Regeln schon für Gesetze der Bastardbildung ansehen und ohe weitere Kritik auf Hieracium ausdehnen wollte. Gelingt es auf dem Wege des Experimentes eine genügende Einsicht in die Bastardbildung der Hieracien zu erhalten, dann wird mit Zuhilfenahme der Erfahrungen, welche über die Vegetationsverhältnisse der verschiedenen wild wachsenden Pflanzen gesammelt wurde, ein competentes Urtheil in dieser Frage möglich werden.
Damit ist zugleich der Zweck ausgesprochen, den die in Rede stehenden Versuche anstreben. Ich erlaube mir nun mit Berücksichtigung dieses Zweckes die bisherigen noch sehr geringen Ergebnisse kurz zusammenzufassen.
1. Bezüglich der Gestalt der Bastarde haben wir die auffallende Erscheinung zu registriren, dass die bis jetzt aus gleicher Befruchtung erhaltenen Formen nicht identisch sind. Die Bastarde H. praealtum + H aurantiacum und H. Auricula + H. aurantiacum sind durch je zwei, H. Auricula und H. pratense ist durch drei Exemplare vertreten, während von den übrigen bisher nur je eines erhalten wurde. Wenn wir die einzelnen Merkmale dieser Bastarde mit den correspondierenden Charakteren der beiden Stammeltern vergleichen, so finden wir, dass dieselben theils Mittelbildungen darstellen, theils aber dem einen der beiden Stamm-Merkmale so nahe stehen, dass das andere weit zurücktritt oder fast der Beobachtung entschwindet. So z. B. sehen wir an der einen der beiden Formen von H. Auricula + H. aurantiacum rein gelbe Scheibenblüthen, nur die Ligeln der Randblümchen sind an der Aussenseite kaum merklich roth angehaucht; bei der anderen hingegen kommt die Blüthenfarbe jener des H. aurantiacum sehr nahe, nur gegen die Mitte der Scheibe hin geht das Orangeroth in ein sattes Goldgelb über. Dieser Unterschied ist beachtenswerth, da die Blüthenfarbe bei Hieracien die Geltung eines constanten Merkmales besitzt. Anderen ähnliche Fälle finden sich an den Blättern, Blüthenständen u. s. w.
Vergleicht man die Bastarde mit den Stammeltern nach der Gesammtheit ihrer Merkmale, dann stellen die beiden Formen des H. praealtum + H. aurantiacum nahezu Mittelformen dar, die jedoch in einzelnen Merkmalen nicht übereinstimmen. Dagegen sehen wir bei H. Auricula + H. aurantiacum und H. Auricula + H. pratense die Formen weit auseinandergehen, so zwar, dass eine davon sich der einen, die andere der zweiten Stammpflanze nahe stellt, während bei dem zuletzt genannten Bastarde noch eine dritte vorhanden ist, welche zwischen beiden fast die Mitte hält.
Es drängt sich von selbst die Vermuthung auf, dass wir hier nur einzelne Gliedre aus noch unbekannten Reihen vor uns haben, welche durch die unmittelbare Einwirkung des Pollens der einen Art auf die Keimzellen einer andren gebildet werden.
2. Die besprochenen Bastarde bilden, mit Ausnahme eines einzigen, keimfähige Samen. Als vollkommen fruchtbar ist zu bezeichnen: H. echioides + H. aurantiacum, als fruchtbar H. praealtum + H. flagellare, als theilweise fruchtbar H. praealtum + H. aurantiacum und H. Auricula + H. pratense, als wenig fruchtbar H. Auricula + H. Pilosella, als unfruchtbar H. Auricula und H. aurantiacum. Von den beiden Formen des zuletzt genannten Bastardes war die roth blühende ganz steril, von der gelb blühenden wurde ein einziger gut ausgebildeter Same erhalten. Ferner kann nicht unerwähnt bleiben, dass unter den Sämlingen des theilweise fruchtbaren Bastardes H. praealtum + H. aurantiacum eine Pflanze die vollkommene Fruchtbarkeit erlangt hat.
Die aus Selbstbefruchtung hervorgegangenen Nachkommen der Bastarde haben bis jetzt nicht variirt, sie stimmen in ihren Merkmalen untereinander und mit der Bastardpflanze, von welcher sie abstammen, überein. Von H. praealtum + H. flagellare sind bis jetzt 2 Generationen, von H. echioides + H. aurantiacum, H. praealtum + H. aurantiacum, H. Auricula + H. Pilosella je eine Generation in 14 bis 112 Exemplaren zur Blüthe gelangt.
4. Es ist die Thatsache zu constatiren, dass bei dem vollkommen fruchtbaren Bastarde E. echioides + H. aurantiacum der Pollen der Stammeltern nicht im Stande war, die Selbstbefruchtung zu hindern, obwohl derselbe den Narben, während sie beim Aufblühen der Antherenröhrchen hervortraten, in grosser Menge mitgetheilt wurde.
Aus zwei auf diese Weise behandelten Blüthenköpfchen wurden durchaus mit der Bastardpflanze übereinstimmende Sämlinge erhalten. Ein ganz ähnlicher Versuch, der schon im heurigen Sommer an dem theilweise fruchtbaren Bastarde H. praealtum + H. aurantiacum vorgenommen wurde, hat zu dem Ergebnisse geführt, dass jene Blüthenköpfchen, an welchen die Narben mit dem Pollen der Stammeltern oder anderer Arten belegt wurden, eine merklich grössere Anzahl guter Samen entwickelten, als jene, welche der Selbstbefruchtung überlassen blieben. Die Erklärung dieser Erscheinung dürfte bei dem Umstande, dass ein grosser Theil der Pollenkörper des Bastardes unter dem Mikroskope eine mangelhafte Ausbildung zeigt, wohl nur darum zu suchen sein, dass bei dem natürlichen Verlaufe der Selbstbefruchtung ein Theil der conceptionsfähigen Eichen wegen schlechter Beschaffenheit des eigenen Pollens nicht befruchtet wird.
Auch bei wild wachsenden, ganz fruchtbaren Arten kommt es nicht selten vor, dass in einzelnen Blüthenköpfchen die Pollenbildung fehlschlägt und in mancher Anthere auch nicht ein einziges gutes Körnchen entwickelt wird. Wenn in solchen Fällen dennoch Samen gebildet werden, so muss die Befruchtung durch fremde Pollen erfolgt sein. Dabei können leicht Bastarde entstehen, indem mancherlie Insekten, namentlich geschäftige Hymenopteren, die Hieracium-Blüthen mit grosser Vorliebe besuchen und sicherlich dafür Sorge tragen, dass der an ihrem haarigen Körper leicht anhängende Pollen benachbarter Pflanzen auf die Narben gelangt.
Aus dem Wenigen, das ich hier mittheilen kann, wird ersichtlich, dass die Arbeit kaum noch über ihre ersten Anfänge hinausreicht. Ich musste wohl Bedenken tragen, an diesem Orte eben erst begonnene Versuche zu besprechen. Nur die Überzeugung, dass die Durchführung der projectirten Experimente noch eine Reihe von Jahren in Anspruch nehmen müsse, und die Ungewißheit, ob es mir vergönnt sein wird, dieselben zu Ende zu führen, konten mich zu der heutigen Mittheilung bestimmen. Durch die Güte des Herrn Directors Dr. Nägeli in München, welcher mir fehlende Arten, namentlich aus den Alpen freundlichst zugesendet hat, bin ich nun in den Stand gesetzt, eine grössere Anzahl von Formen in den Kreis der Versuche zu ziehen und darf hoffen, schon im kommenden Jahre Einiges zur Ergänzung und Sicherstellung der heutigen Angaben nachholen zu können.
Wenn wir schliesslich die besprochenen, allerdings noch sehr unsicheren Resultate mit jenen vergleichen, welche aus Kreuzungen im Jahre 1865 hier mitzutheilen die Ehre hatte, so begegnen wir einer sehr wesentlichen Verschiedenheit. Bei Pisum haben die Bastarde, welche unmittelbar aus der Kreuzung zweier Formen gewonnen werden, in allen Fällen den gleichen Typus, ihre Nachkommen dagegen sind veränderlich und variiren nach einem bestimmten Gesetze. Bei Hieracium scheint sich nach den bisherigen Versuchen das gerade Gegentheil davon herausstellen zu wollen. Schon bei Besprechung der Pisum-Versuche wurde darauf hingewiesen, dass es auch Bastarde gibt, deren Nachkommen nicht variiren, dass z. B. nach Wichura die Bastarde von Salix sich unverändert wie reine Arten fortpflanzen. Wir hätten demnach bei Hieracium eine analogen Fall. Ob man bei diesem Umstande die Vermuthung aussprechen dürfe, dass die Polymorphie der Gattungen Salix und Hieracium mit dem eigentlichen Verhalten ihrer Bastarde in Zusammenhang stehe, das ist bis jetzt noch eine Frage, die sich wohl anregen, nicht aber beantworten lässt.
|