
Die DNS ist im Zellkern mit Histonen komplexiert und formt damit das
sogenannte Chromatin. In den meisten eukaryotischen Zellen kommen fünf
verschiedene Histone vor: H1, H2a, H2b, H3, H4. Ein Aggregat aus je zwei
Molekülen H2a, H2b, H3 und H4 bildet ein Nukleosom, eine scheibenförmige
Struktur, um die DNS herumgewickelt ist, und aus der Teile der Histonmoleküle nach außen ragen. Auf je ca. 200 Basenpaare entfällt
ein Nukleosom, 140 der Basenpaare sind in 1 3/4 Windungen um den Histonkomplex
gewunden, die übrigen 60 sind frei und verbinden Nukleosomen untereinander.
Das H1 gehört nicht dem Nukleosomenkomplex an, sondern dient dazu,
das Chromatin noch weiter zu kondensieren.
 |
|
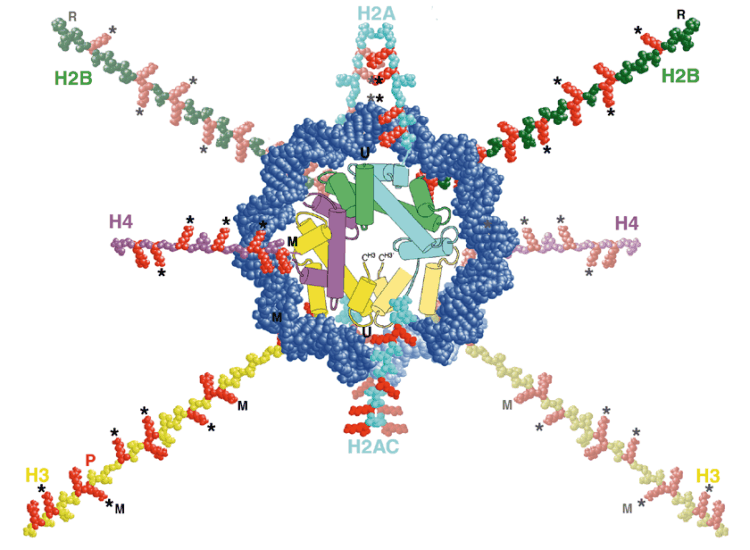
Nukleosomenmodell: Aus: A. P. WOLFFE & J. J. HAYES - Chromatin disruption and modification. - Nucleic Acid Research, 27, 711-720 (1999)
Es gibt Anhaltspunkte dafür, daß bestimmte Proteinfraktionen an bestimmte DNS-Abschnitte binden; es gibt ferner experimentelle Beweise dafür, daß sie durch Hormone, Metaboliten u.a. aktiviert werden können, und es gibt schließlich Hinweise darauf, daß über das Phytochromsystem Signale an die DNS weitergegeben werden, die selektiv die Transkription bestimmter Abschnitte fördern.
Zu den sauren Kernproteinen zählen auch die Polymerasen, die die Replikation und Transkription katalysieren, sowie Proteine, die mit der RNS RNP-Komplexe (Ribonukleoproteinkomplexe) bilden.
Der Grad der Kondensation der DNS im Kern ist variabel. Stark kondensierte DNS wird als Heterochromatin, weniger stark kondensierte als Euchromatin bezeichnet. Heterochromatin ist durch bestimmte Kernfarbstoffe intensiver anfärbbar als das Euchromatin. An anderer Stelle wird eine cytologische Definition der beiden Begriffe gegeben, und es wird deutlich gemacht, daß sie zwar nützlich und brauchbar ist, aber keine klaren Entweder-oder-Zustände beschreibt.
Kondensation und Dekondensation der DNS sind, zum Teil wenigstens, reversible Prozesse. Jeder DNS-Abschnitt, der repliziert oder transkribiert wird, muß zumindest während dieser Zeiträume dekondensiert sein, wohingegen die DNS während der Zellteilung in den Chromosomen (Mitose, Meiose) den höchsten Kondensationsgrad erreicht.
Wie schon erwähnt, besteht eukaryotische DNS aus repetitiven (r) und nichtrepetitiven, singulären (s) Abschnitten, die in drei verschiedenen Organisationsmustern auftreten.
Die repetitiven Abschnitte (Sequenzen) sind an einer oder an wenigen Stellen im Genom konzentriert. Sie bilden Folgen wie
rlrlr1 . . .r2r2r2r2. . .r3r3r3........rnrnrn
Die Zahl der verschiedenen Repetitionseinheiten (rn) und die Zahl der Wiederholungen liegt in Größenordnungen von weit über 1000. In der englischen Literatur spricht man von Clustern.
Die repetitiven Sequenzen können in Gruppen mit singulären wechseln (interspersed pattern)
r1r1r1 . . .r2r2. . .s1 . . .r3r3r3. . .s2s3s4r4r4r4. . .rn
In bestimmten Teilen der DNS findet man Abschnitte wie die unter (2) beschriebenen, in anderen vorwiegend solche, die nur singuläre Sequenzen enthalten oder in denen singuläre Sequenzen von wenigen repetitiven unterbrochen sind.

Teile von Lampenbürstenchromosomen von Triturus cristatus (einer amerikanischen Molchart): "intermediär" repetitive DNS wurde in situ gegen [3H]- markierte RNS-Transkripte an den Schlaufen der Lampenbürstenchromosomen hybridisiert. Nur einige wenige Schlaufenpaare sind auf diese Weise autoradiographisch markiert (H. C. MacGREGOR, Leicester, 1977)
|
|