

Part 3
Coals mainly occur in the Westphalian. However, at the end of the Westphalian the considerable changes in the composition of the flora can be seen. The hitherto widespread tree-like lycophytes largely became extinct; only very few forms locally persist into the Stephanian. In the Rotliegend (Early Permian) of Europe and North America only a single tree-like lycopsid is known: Sigillaria brardii. This plant was very tolerant with regard to its habitat, although it always grew in humid environments.
Although this floral change has thoroughly been documented in recent years, also by palynological data (Phillips et al. 1985), the causes have never been explained in a satisfactory way. One of the problems is that there are hardly successions in which the Westphalian-Stephanian transition is developed without hiatuses. A further problem is that the Westphalian and Stephanian have been defined in different types of basins; the Westphalian in a so-called paralic basin and the Stephanian in an intramontane basin. Floras in such fundamentally different types of basins are not necessarily well comparable. Virtually all our Stephanian floras are known from intramontane basins.
Several authors relate the almost complete extinction of arborescent lycopods to climatic changes (e.g., DiMichele et al. 1992). From the Westphalian onwards northwest and central Europe drifted in northward direction. In the Westphalian this area was still in an equatorial position, by the end of the Permian it had reached a position of approx. 30° N. This means that the continental plates wandered through the climatic zones. Although this is certainly an important factor, also tectonic processes have recently been discussed in relation to this floral change (Kerp 1996).
|
During the Namurian and the Westphalian the subsidence of the
depositional basins was more or less equal to the sedimentation
rate. Therefore thick peat deposits could develop over large
areas. These peat deposits later became coal. Only occasionally
transgressions occurred, which, however, could lead to the
flooding of large areas. In the mainly terrestrial succession of
the Namurian and Westphalian marine horizons occur, some of which
even have been traced over a area of 600,000 km². However, these
rather short-termed floodings have had little effect on the
flora. Most plant species could apparently retreat to refugia
from which they could migrate back after the lowering of the sea
level. In the Westphalian the marine influences diminished and
the youngest marine intercalations are known from the Stephanian.
By the end of the Westphalian the northward moving southern, so-called Gondwana continental plate collided with the Eurasian (or Laurussian) plate (Ziegler 1990). As a result of this collision sediments were folded up and the Variscan Mountain Chain was formed (Fig. 4). This also resulted a strong reduction of the Paralic Coal Basin which was situated immediately north of the folded area; only a few small relict basins were left. The sediments in these small basins express much higher sedimentation dynamics. The folding and formation of the Variscan Mountain Chain had resulted in a stronger erosion and the sediments of the uppermost Westphalian and Stephanian are therefore generally sandier than the underlying Westphalian deposits. In such sandy sediments the ground water level will have been lower than in the predominantly clayey sediments of the Westphalian, in which stagnation of the ground and rain water locally resulted in the formation of suitable biotopes for arborescent lycophytes. The development of the Variscan Mountain Chain will certainly also have influenced atmospheric circulation patterns. Moreover, it should also be realised that the reduction of a once widespread wetland area will have had some effect. The replacement of an extended wetland area with a luxurious vegetation, a more or less self-regulating ecosystem with a considerable water-storage capacity, by a mountainous area will have lead to changes in atmospherical humidity. Arborescent lycophytes and other plants strongly adapted to very stable ecological conditions could apparently not persist under the higher sedimentation and climate dynamics during the Stephanian and became extinct. It is clear that there is not a single reason for the extinction of the arborescent lycophytes, but that it is the result of a complex many, often interrelated factors. |
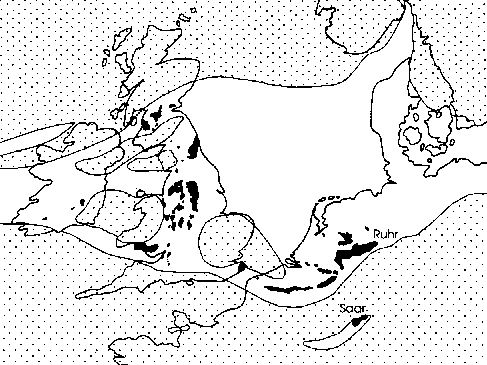 Abb. 4a | |||
 Abb. 4b Abb. 4: Palaeogeographical reconstructions of the Westfalian and Stefanian. Modified after Ziegler (1990). | ||||
 |
terrestrial and nearshore marine deposits |  |
Variscan fold belt | |
 |
Land areas |  |
Front of the Variscan fold belt with overthrust | |
 |
Volcanism |  |
Coal deposits | |
Although the Stephanian is still partly coal-bearing, in the Rotliegend a few thin coal seams are only very locally developed. In practice it appears to be very difficult to define a clear boundary between the Stephanian and Rotliegend because several plant taxa which have previously been regarded as marker fossils now appear to be strongly facies-dependant. These therefore rather reflect different ecological conditions than that they indicate different ages. The biostratigraphical value of these fossils is thus very limited. Another major problem is the facies development in the individual basins. Especially the Rotliegend is characterised by many rapid lateral and horizontal changes in facies; this can also have influenced the composition of the floras.
The individual, mostly rather small intramontane basins, especially the Rotliegend basins, are often very difficult to correlate with each other. In some cases such correlations are also strongly hampered by the presence of considerable hiatuses. Stephanian floras are much more common and therefore much better known than Rotliegend floras. Especially in younger deposits plant fossils become rare. Although we know a relatively large number of plant localities from the Lower Rotliegend, there are only very few localities known from the Upper Rotliegend. Floras which are most probably coeval can be developed very differently. In these cases palynology, the study of fossil spores and pollen does not help any further, as palynomorphs are usually absent or very poorly preserved in redbed sequences.
 Fig. 5: A conifer from the Rotliegend of the Saar-Nahe Basin. |
The flora of the Stephanian can be characterised by a sudden decrease of arborescent lycophytes and a dominance of tree ferns; also seed ferns and cordaites are well represented. During the Stephanian the conifers, a group of gymnosperms whose earliest representatives looked very similar to modern araucarians, became gradually more common. The earliest conifers are known from the Middle Westphalian, but these records consist of small fragments which have apparently been transported over considerable distances. The increase in dominance of conifers continues in the Rotliegend, although completely conifer-dominated floras are rare in the Rotliegend. Other new elements are the more modern types of pteridosperms, like the callipterids. Since the Stephanian a reduction of the number of species can be seen. |
As early as 1930 Gothan & Gimm noticed the existence of different types of floral associations in the European Rotliegend. These are the peat-forming floras which indicate humid biotopes and the hinterland floras with elements which grew under drier conditions. Detailed studies have resulted in the recognition of a larger number of other typical associations (e.g., Barthel 1976). The hinterland or extra-basinal floras are usually poorly and incompletely preserved as these plants grew outside the sedimentation basins and the material was transported for a longer distance before it was embedded in the sediment. The preservation potential of such floral associations is therefore much lower than that of those growing in basinal areas.
Although many Rotliegend floras predominantly consist of hygro- and hydrophilous elements, the number of taxa adapted to drier conditions generally increases in younger deposits. The hygro- and hydrophilous floras of the Rotliegend have a rather archaic character and include many long-ranging, essentially Stephanian taxa. Evolutionary innovations, especially within the gymnosperms, took place in the extra-basinal areas. Several groups which first appeared during the Stephanian and Rotliegend, such as Cycadophytes and Ginkgophytes consist of typical hinterland elements. It is very difficult, if not impossible, to reconstruct the development of hinterland floras because of the paucity of the fossil record.
Although the general picture is more or less clear, many detailed questions on Stephanian and Rotliegend floras still remain unanswered. Anyhow, it is obvious that there was a clear differentiation in basinal and hinterland floras. The first are characterised by the dominance of tree ferns such as Psaronius in the petrified forest of Chemnitz (Rößler 1996); the latter were dominated by conifers.